

Distinct cell cycle-dependent roles for dynactin and dynein at centrosomes
- PMID: 12391026
- PMCID: PMC2173046
- DOI: 10.1083/jcb.200203089
Distinct cell cycle-dependent roles for dynactin and dynein at centrosomes
Abstract
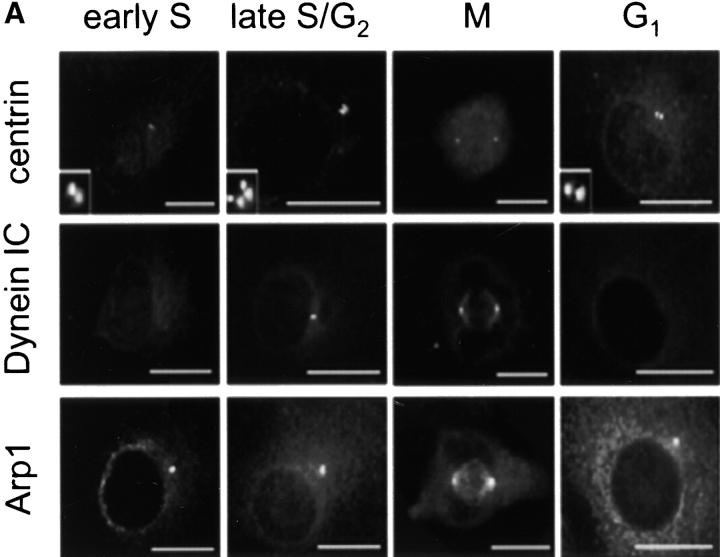
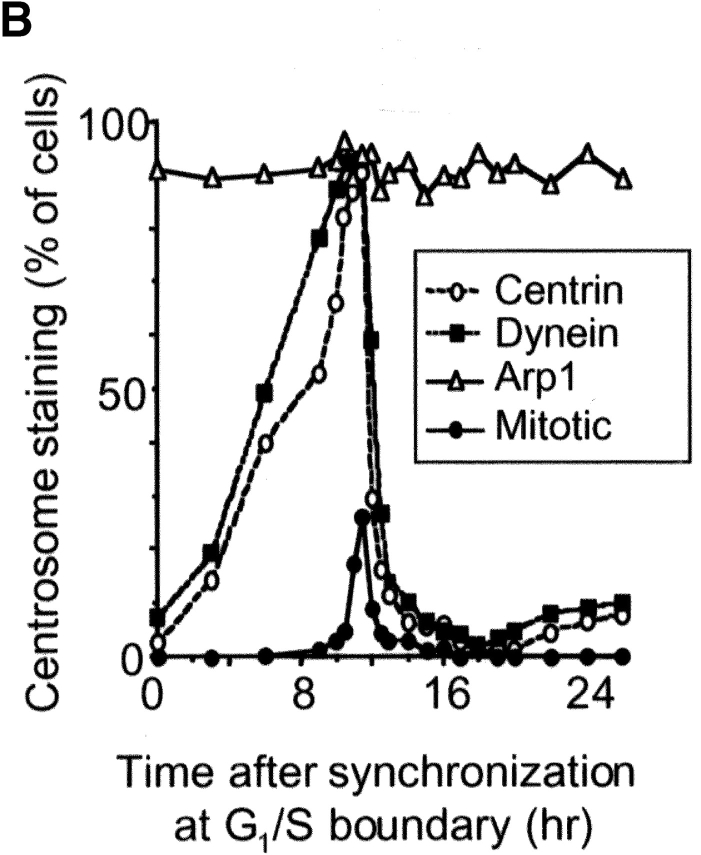
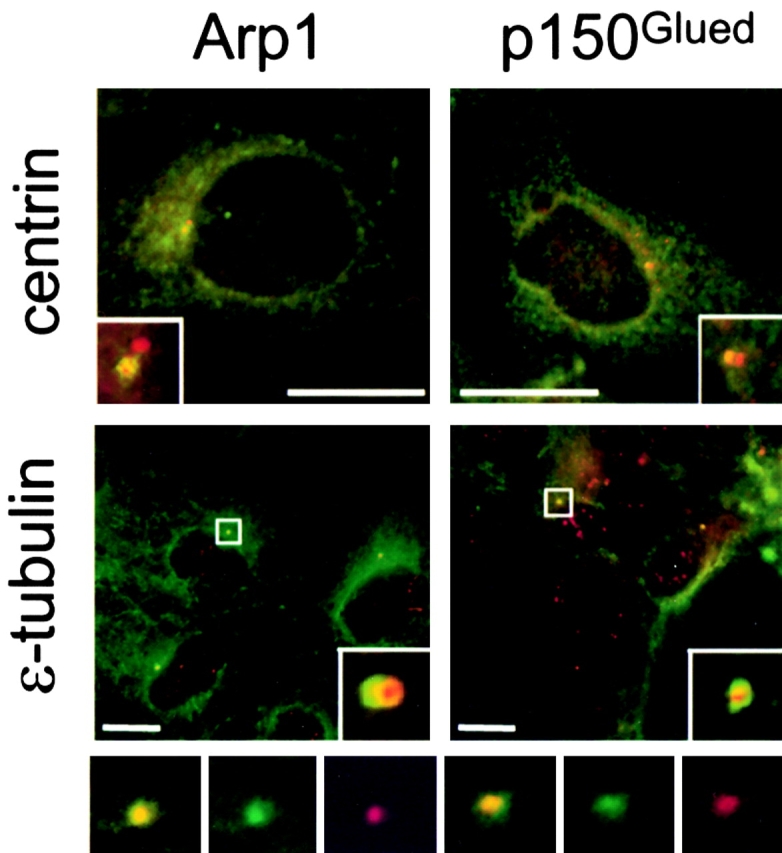
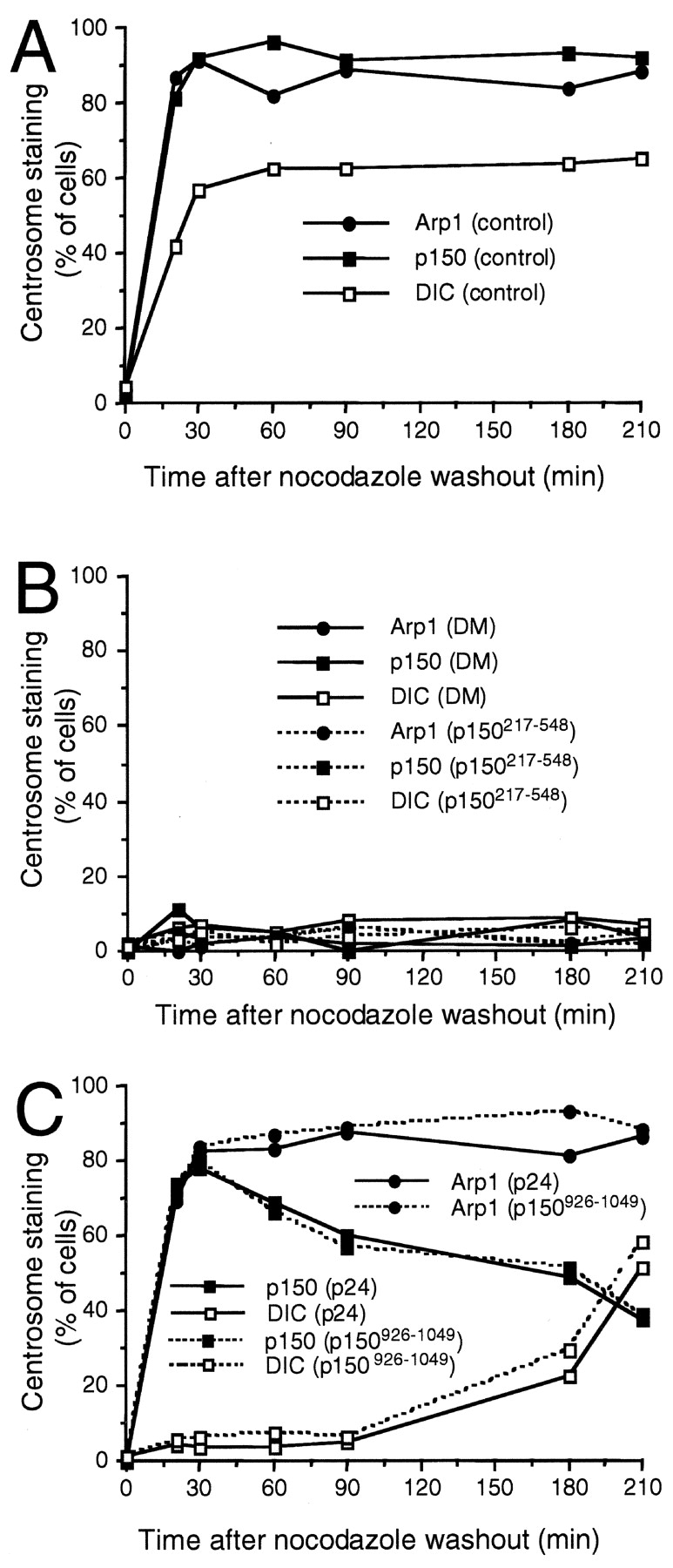
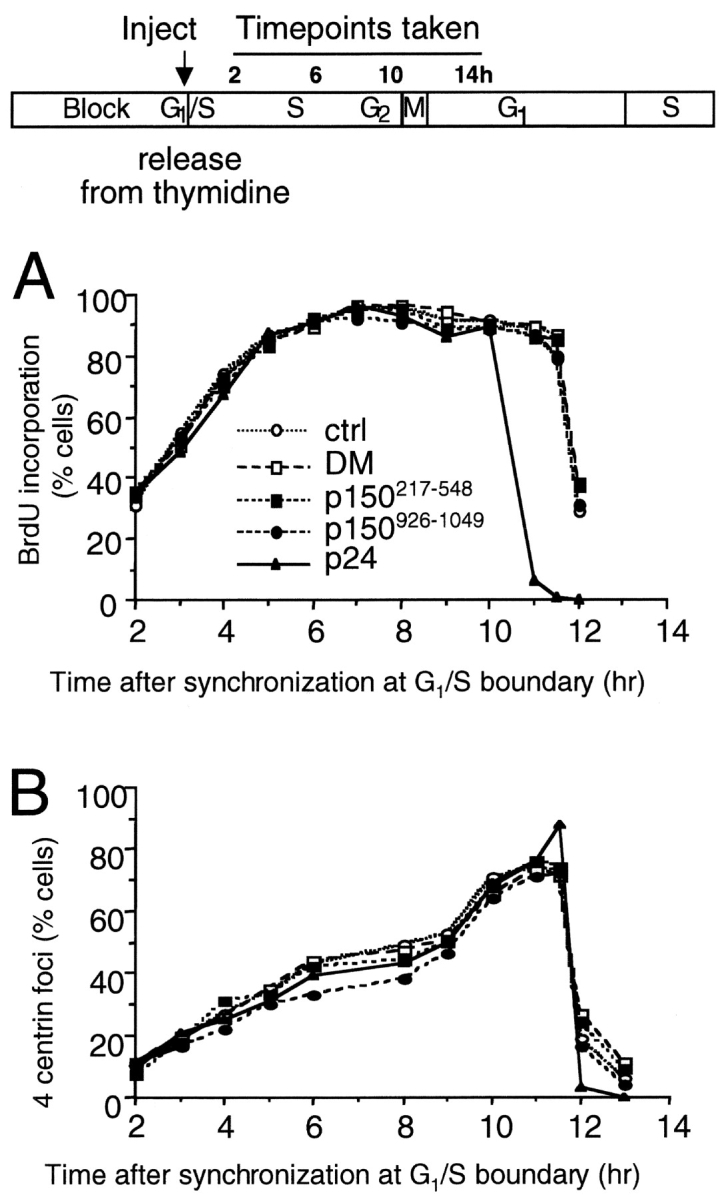
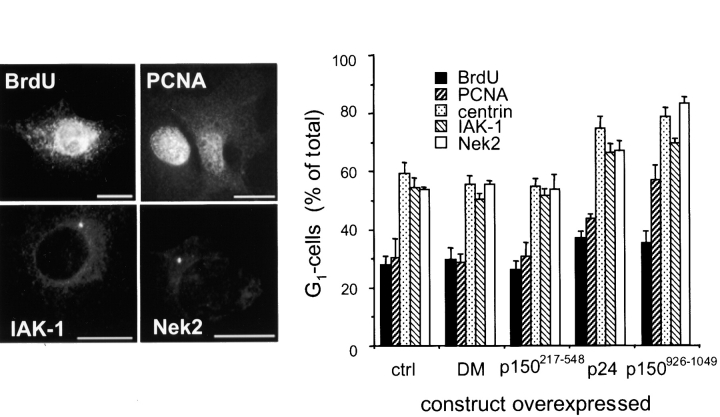
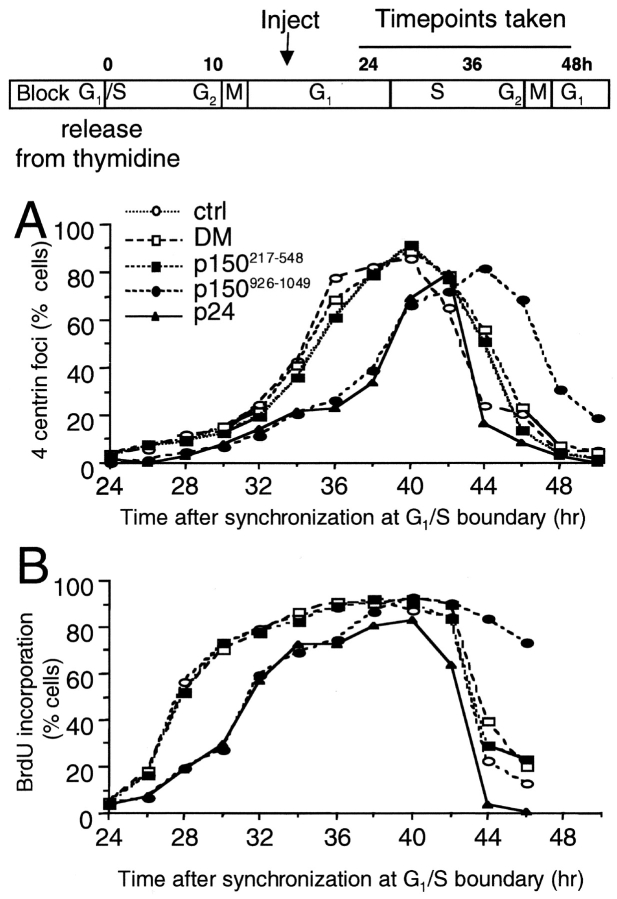
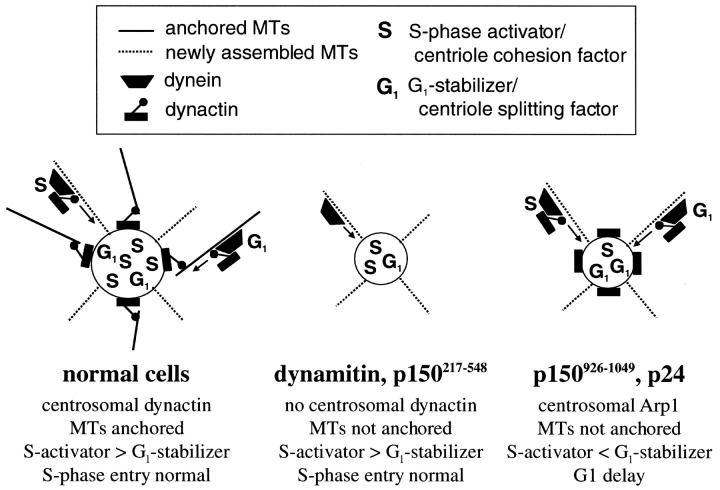
Centrosomal dynactin is required for normal microtubule anchoring and/or focusing independently of dynein. Dynactin is present at centrosomes throughout interphase, but dynein accumulates only during S and G2 phases. Blocking dynein-based motility prevents recruitment of dynactin and dynein to centrosomes and destabilizes both centrosomes and the microtubule array, interfering with cell cycle progression during mitosis. Destabilization of the centrosomal pool of dynactin does not inhibit dynein-based motility or dynein recruitment to centrosomes, but instead causes abnormal G1 centriole separation and delayed entry into S phase. The correct balance of centrosome-associated dynactin subunits is apparently important for satisfaction of the cell cycle mechanism that monitors centrosome integrity before centrosome duplication and ultimately governs the G1 to S transition. Our results suggest that, in addition to functioning as a microtubule anchor, dynactin contributes to the recruitment of important cell cycle regulators to centrosomes.
Figures
References
-
- Allan, V. 2000. Dynactin. Curr. Biol. 10:R432. - PubMed
-
- Bornens, M. 2002. Centrosome composition and microtubule anchoring mechanisms. Curr. Opin. Cell Biol. 14:25–34. - PubMed
-
- Bouckson-Castaing, V., M. Moudjou, D.J. Ferguson, S. Mucklow, Y. Belkaid, G. Milon, and P.R. Crocker. 1996. Molecular characterisation of ninein, a new coiled-coil protein of the centrosome. J. Cell Sci. 109:179–190. - PubMed
-
- Bravo, R., and J.E. Celis. 1985. Changes in the nuclear distribution of cyclin (PCNA) during S-phase are not triggered by post-translational modifications that are expected to moderately affect its charge. FEBS Lett. 182:435–440. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
