

Oct-1 potentiates CREB-driven cyclin D1 promoter activation via a phospho-CREB- and CREB binding protein-independent mechanism
- PMID: 12391146
- PMCID: PMC134723
- DOI: 10.1128/MCB.22.22.7769-7779.2002
Oct-1 potentiates CREB-driven cyclin D1 promoter activation via a phospho-CREB- and CREB binding protein-independent mechanism
Abstract
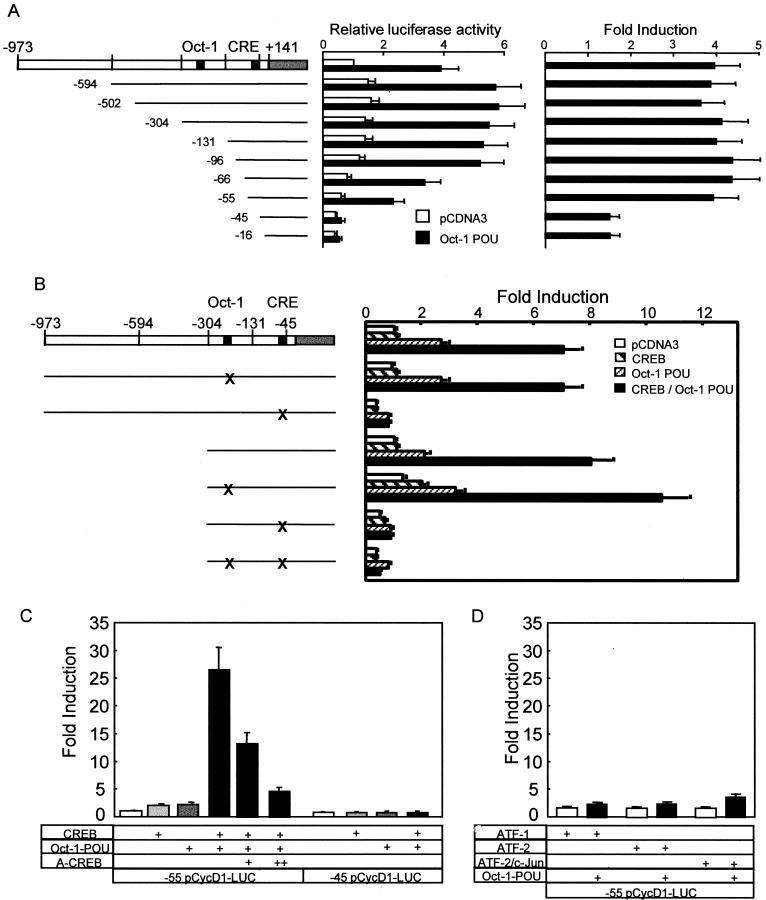
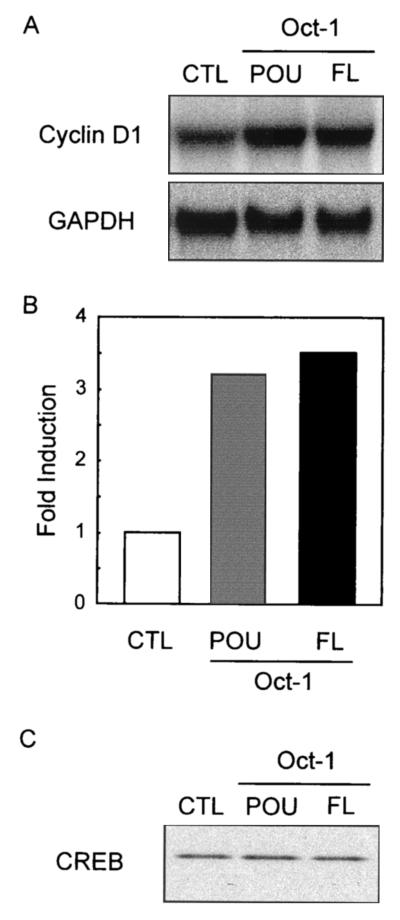
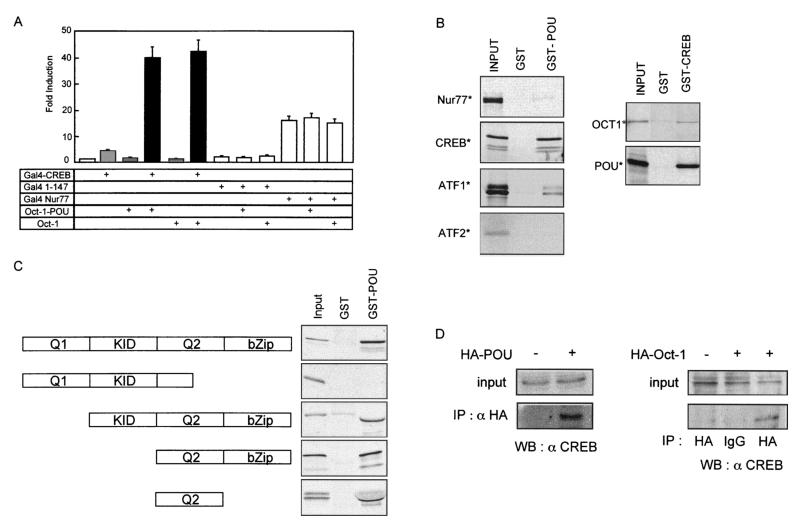
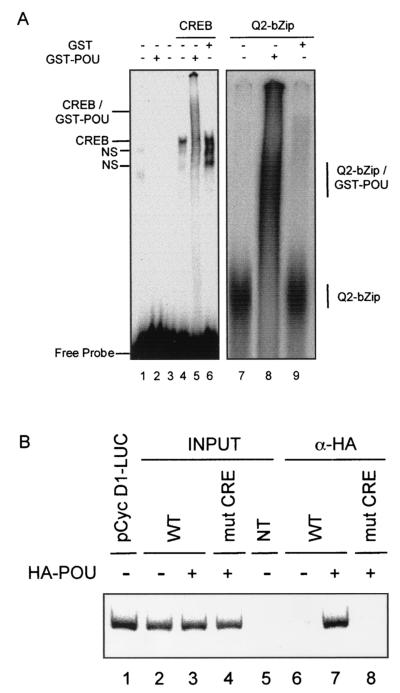
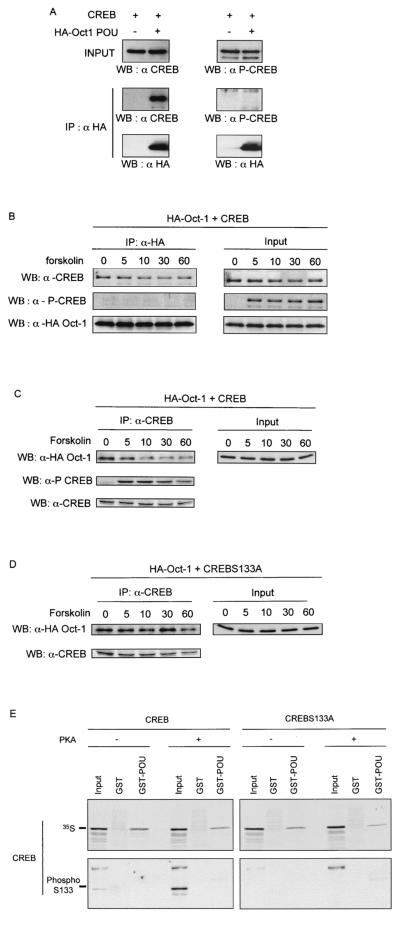
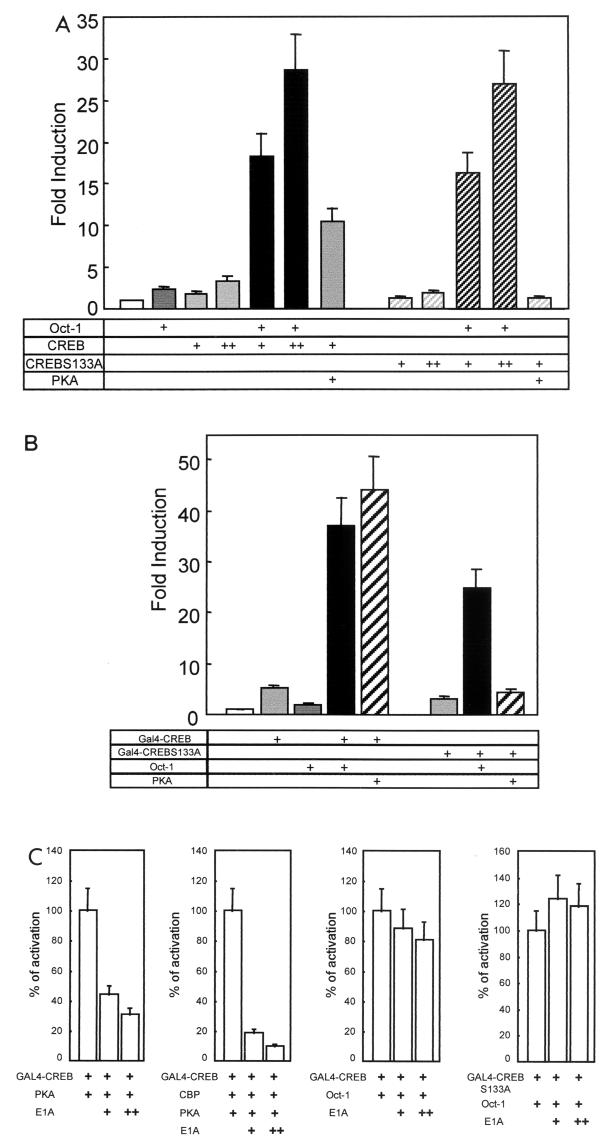
Cyclin D1, the regulatory subunit for mid-G(1) cyclin-dependent kinases, controls the expression of numerous cell cycle genes. A cyclic AMP-responsive element (CRE), located upstream of the cyclin D1 mRNA start site, integrates mitogenic signals that target the CRE-binding factor CREB, which can recruit the transcriptional coactivator CREB-binding protein (CBP). We describe an alternative mechanism for CREB-driven cyclin D1 induction that involves the ubiquitous POU domain protein Oct-1. In the breast cancer cell line MCF-7, overexpression of Oct-1 or its POU domain strongly increases transcriptional activation of cyclin D1 and GAL4 reporter genes that is specifically dependent upon CREB but independent of Oct-1 DNA binding. Gel retardation and chromatin immunoprecipitation assays confirm that POU forms a complex with CREB bound to the cyclin D1 CRE. In solution, CREB interaction with POU requires the CREB Q2 domain and, notably, occurs with CREB that is not phosphorylated on Ser 133. Accordingly, Oct-1 also potently enhances transcriptional activation mediated by a Ser133Ala CREB mutant. Oct-1/CREB synergy is not diminished by the adenovirus E1A 12S protein, a repressor of CBP coactivator function. In contrast, E1A strongly represses CBP-enhanced transactivation by CREB phosphorylated on Ser 133. Our observation that Oct-1 potentiates CREB-dependent cyclin D1 transcriptional activity independently of Ser 133 phosphorylation and E1A-sensitive coactivator function offers a new paradigm for the regulation of cyclin D1 induction by proliferative signals.
Figures
Similar articles
-
Chromatin-dependent cooperativity between constitutive and inducible activation domains in CREB.Mol Cell Biol. 2001 Dec;21(23):7892-900. doi: 10.1128/MCB.21.23.7892-7900.2001. Mol Cell Biol. 2001. PMID: 11689682 Free PMC article.
-
Activation of the cyclin D1 gene by the E1A-associated protein p300 through AP-1 inhibits cellular apoptosis.J Biol Chem. 1999 Nov 26;274(48):34186-95. doi: 10.1074/jbc.274.48.34186. J Biol Chem. 1999. PMID: 10567390
-
STAT5 and Oct-1 form a stable complex that modulates cyclin D1 expression.Mol Cell Biol. 2003 Dec;23(24):8934-45. doi: 10.1128/MCB.23.24.8934-8945.2003. Mol Cell Biol. 2003. PMID: 14645506 Free PMC article.
-
Regulation of somatostatin gene transcription by cyclic adenosine monophosphate.Metabolism. 1996 Aug;45(8 Suppl 1):4-7. doi: 10.1016/s0026-0495(96)90068-2. Metabolism. 1996. PMID: 8769368 Review.
-
The regulation of cyclin D promoters - review.Cancer Treat Res Commun. 2021;27:100338. doi: 10.1016/j.ctarc.2021.100338. Epub 2021 Feb 17. Cancer Treat Res Commun. 2021. PMID: 33618151 Review.
Cited by
-
Selective CREB-dependent cyclin expression mediated by the PI3K and MAPK pathways supports glioma cell proliferation.Oncogenesis. 2014 Jun 30;3(6):e108. doi: 10.1038/oncsis.2014.21. Oncogenesis. 2014. PMID: 24979279 Free PMC article.
-
Small molecule inhibition of cAMP response element binding protein in human acute myeloid leukemia cells.Leukemia. 2016 Dec;30(12):2302-2311. doi: 10.1038/leu.2016.139. Epub 2016 May 23. Leukemia. 2016. PMID: 27211267 Free PMC article.
-
The Oct1 transcription factor and epithelial malignancies: Old protein learns new tricks.Biochim Biophys Acta. 2016 Jun;1859(6):792-804. doi: 10.1016/j.bbagrm.2016.02.007. Epub 2016 Feb 10. Biochim Biophys Acta. 2016. PMID: 26877236 Free PMC article. Review.
-
Activation of CREB/ATF sites by polyomavirus large T antigen.J Virol. 2005 Apr;79(7):4180-90. doi: 10.1128/JVI.79.7.4180-4190.2005. J Virol. 2005. PMID: 15767419 Free PMC article.
-
CREB1 contributes colorectal cancer cell plasticity by regulating lncRNA CCAT1 and NF-κB pathways.Sci China Life Sci. 2022 Aug;65(8):1481-1497. doi: 10.1007/s11427-022-2108-x. Epub 2022 Jun 10. Sci China Life Sci. 2022. PMID: 35696016
References
-
- Albanese, C., M. D'Amico, A. T. Reutens, M. Fu, G. Watanabe, R. J. Lee, R. N. Kitsis, B. Henglein, M. Avantaggiati, K. Somasundaram, B. Thimmapaya, and R. G. Pestell. 1999. Activation of the cyclin D1 gene by the E1A-associated protein p300 through AP-1 inhibits cellular apoptosis. J. Biol. Chem. 274:34186-34195. - PubMed
-
- Billon, N., D. Carlisi, M. B. Datto, L. A. van Grunsven, A. Watt, X. F. Wang, and B. B. Rudkin. 1999. Cooperation of Sp1 and p300 in the induction of the CDK inhibitor p21WAF1/CIP1 during NGF-mediated neuronal differentiation. Oncogene 18:2872-2882. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials