

Transcription through intergenic chromosomal memory elements of the Drosophila bithorax complex correlates with an epigenetic switch
- PMID: 12391168
- PMCID: PMC134728
- DOI: 10.1128/MCB.22.22.8026-8034.2002
Transcription through intergenic chromosomal memory elements of the Drosophila bithorax complex correlates with an epigenetic switch
Abstract
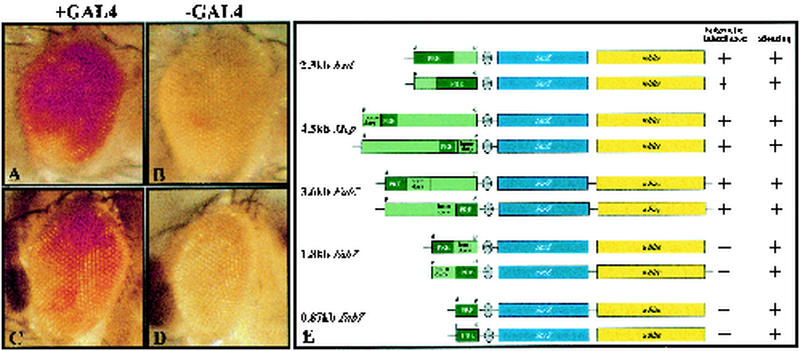
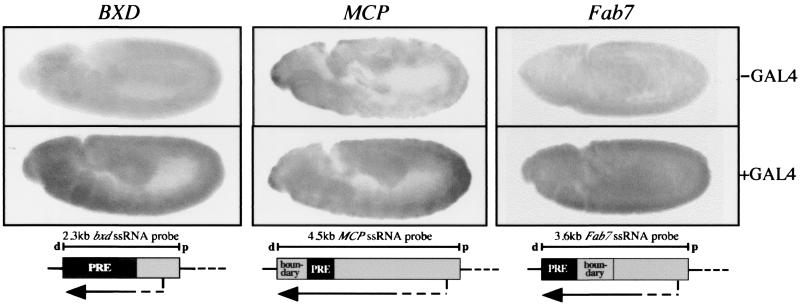
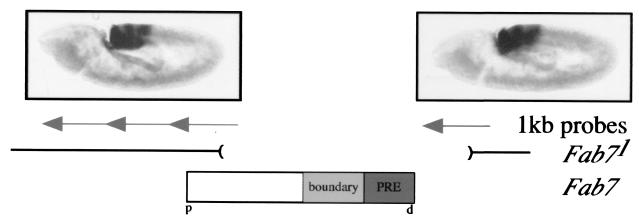
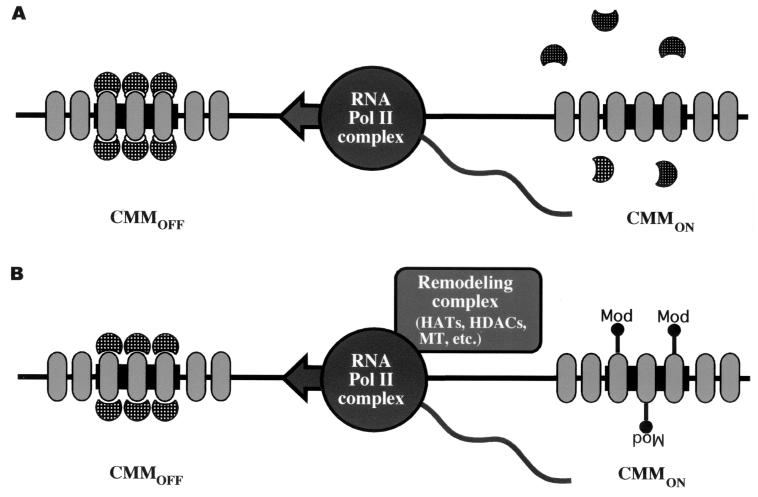
The proteins of the trithorax and Polycomb groups maintain the differential expression pattern of homeotic genes established by the early embryonic patterning system during development. These proteins generate stable and heritable chromatin structures by acting via particular chromosomal memory elements. We established a transgenic assay system showing that the Polycomb group response elements bxd and Mcp confer epigenetic inheritance throughout development. With previously published data for the Fab7 cellular memory module, we confirmed the cellular memory function of Polycomb group response elements. In Drosophila melanogaster, several of these memory elements are located in the large intergenic regulatory regions of the homeotic bithorax complex. Using a transgene assay, we showed that transcription through a memory element correlated with the relief of silencing imposed by the Polycomb group proteins and established an epigenetically heritable active chromatin mode. A memory element remodeled by the process of transcription was able to maintain active expression of a reporter gene throughout development. Thus, transcription appears to reset and change epigenetic marks at chromosomal memory elements regulated by the Polycomb and trithorax proteins. Interestingly, in the bithorax complex of D. melanogaster, the segment-specific expression of noncoding intergenic transcripts during embryogenesis seems to fulfill this switching role for memory elements regulating the homeotic genes.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous