

Tissue-specific RNA interference in postimplantation mouse embryos with endoribonuclease-prepared short interfering RNA
- PMID: 12391321
- PMCID: PMC137867
- DOI: 10.1073/pnas.192559699
Tissue-specific RNA interference in postimplantation mouse embryos with endoribonuclease-prepared short interfering RNA
Abstract
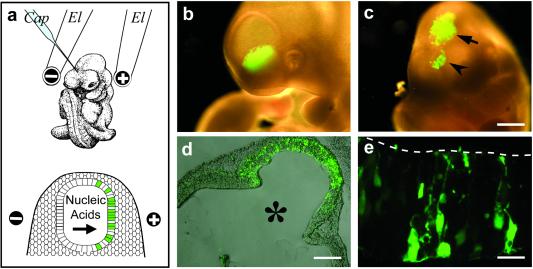
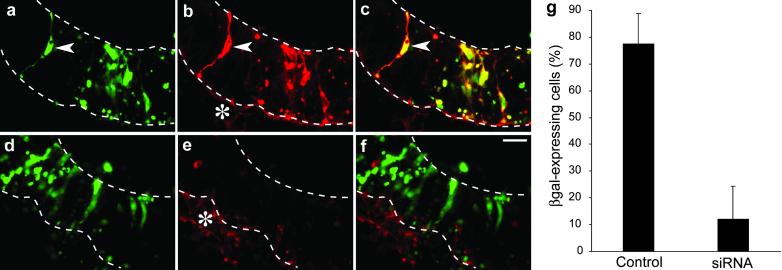
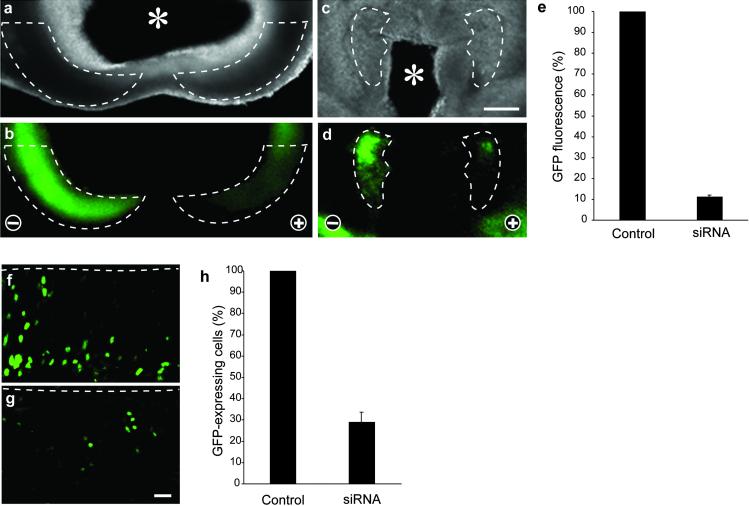
RNA interference (RNAi) using double-stranded RNA has been used for the systematic analysis of gene function in invertebrate organisms. Here we have explored the use of short interfering RNA (siRNA) to knock down gene expression during the development of mammalian postimplantation embryos. The developing CNS system of embryonic day 10 mouse embryos was used as a model tissue. siRNA prepared by endoribonuclease digestion (esiRNA) was injected into the lumen of the neural tube at specific regions and delivered into neuroepithelial cells by directed electroporation. Injected and electroporated embryos were grown for 1 day in whole-embryo culture and the effects of RNAi were examined. esiRNA directed against beta-galactosidase (beta-gal), coelectroporated into neuroepithelial cells together with reporter plasmids expressing GFP and beta-gal, abolished expression of beta-gal but not GFP, showing the specificity of the esiRNA-mediated RNAi. To demonstrate RNAi of endogenous gene expression, we used heterozygous embryos of a knock-in mouse line expressing GFP from the Tis21 locus, a gene turned on in neuroepithelial cells that switch from proliferation to neurogenesis. GFP-directed esiRNA electroporated into neuroepithelial cells of such embryos blocked the GFP expression normally occurring on the onset of neurogenesis. Taken together, our data indicate that esiRNA delivered in a tissue-specific manner by topical injection followed by directed electroporation can efficiently silence endogenous gene expression in mammalian postimplantation embryos.
Figures
Similar articles
-
Tissue-specific RNA interference in post-implantation mouse embryos using directional electroporation and whole embryo culture.Differentiation. 2004 Mar;72(2-3):92-102. doi: 10.1111/j.1432-0436.2004.07202002.x. Differentiation. 2004. PMID: 15066189 Review.
-
Sequence-specific gene silencing in murine muscle induced by electroporation-mediated transfer of short interfering RNA.J Gene Med. 2004 Jan;6(1):105-10. doi: 10.1002/jgm.456. J Gene Med. 2004. PMID: 14716682
-
Spatial and temporal 'knock down' of gene expression by electroporation of double-stranded RNA and morpholinos into early postimplantation mouse embryos.Mech Dev. 2002 Oct;118(1-2):57-63. doi: 10.1016/s0925-4773(02)00191-0. Mech Dev. 2002. PMID: 12351170
-
Efficient RNA interference in zebrafish embryos using siRNA synthesized with SP6 RNA polymerase.Dev Growth Differ. 2005 Jun;47(5):323-31. doi: 10.1111/j.1440-169X.2005.00807.x. Dev Growth Differ. 2005. PMID: 16026540
-
Small RNA: can RNA interference be exploited for therapy?Lancet. 2003 Oct 25;362(9393):1401-3. doi: 10.1016/S0140-6736(03)14637-5. Lancet. 2003. PMID: 14585643 Review.
Cited by
-
MicroRNA-34/449 controls mitotic spindle orientation during mammalian cortex development.EMBO J. 2016 Nov 15;35(22):2386-2398. doi: 10.15252/embj.201694056. Epub 2016 Oct 5. EMBO J. 2016. PMID: 27707753 Free PMC article.
-
TIS21 (/BTG2/PC3) as a link between ageing and cancer: cell cycle regulator and endogenous cell death molecule.J Cancer Res Clin Oncol. 2006 Jul;132(7):417-26. doi: 10.1007/s00432-006-0080-1. Epub 2006 Feb 3. J Cancer Res Clin Oncol. 2006. PMID: 16456675 Free PMC article. Review.
-
Chemically modified oligonucleotide-increased stability negatively correlates with its efficacy despite efficient electrotransfer.J Membr Biol. 2012 Sep;245(9):565-71. doi: 10.1007/s00232-012-9468-9. Epub 2012 Jul 15. J Membr Biol. 2012. PMID: 22797942
-
Complete, gene-specific siRNA libraries: production and expression in mammalian cells.RNA. 2005 May;11(5):837-46. doi: 10.1261/rna.7285805. RNA. 2005. PMID: 15840823 Free PMC article.
-
High-throughput selection of effective RNAi probes for gene silencing.Genome Res. 2003 Oct;13(10):2333-40. doi: 10.1101/gr.1575003. Genome Res. 2003. PMID: 14525931 Free PMC article.
References
-
- Fire A., Xu, S., Montgomery, M. K., Kostas, S. A., Driver, S. E. & Mello, C. C. (1998) Nature 391, 806-811. - PubMed
-
- Fraser A. G., Kamath, R. S., Zipperlen, P., Martinez-Campos, M., Sohrmann, M. & Ahringer, J. (2000) Nature 408, 325-330. - PubMed
-
- Gönczy P., Echeverri, C., Oegema, K., Coulson, A., Jones, S. J., Copley, R. R., Duperon, J., Oegema, J., Brehm, M., Cassin, E., et al. (2000) Nature 408, 331-336. - PubMed
-
- Hannon G. J. (2002) Nature 418, 244-251. - PubMed
-
- Schmid A., Schindelholz, B. & Zinn, K. (2002) Trends Neurosci. 25, 71-74. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
