

Metabolic flux in both the purine mononucleotide and histidine biosynthetic pathways can influence synthesis of the hydroxymethyl pyrimidine moiety of thiamine in Salmonella enterica
- PMID: 12399482
- PMCID: PMC151968
- DOI: 10.1128/JB.184.22.6130-6137.2002
Metabolic flux in both the purine mononucleotide and histidine biosynthetic pathways can influence synthesis of the hydroxymethyl pyrimidine moiety of thiamine in Salmonella enterica
Abstract
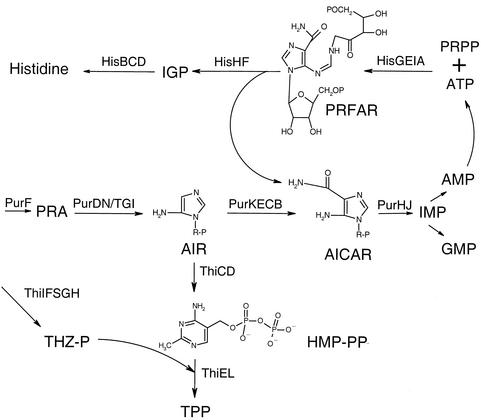
Together, the biosyntheses of histidine, purines, and thiamine pyrophosphate (TPP) contain examples of convergent, divergent, and regulatory pathway integration. Mutations in two purine biosynthetic genes (purI and purH) affect TPP biosynthesis due to flux through the purine and histidine pathways. The molecular genetic characterization of purI mutants and their respective pseudorevertants resulted in the conclusion that <1% of the wild-type activity of the PurI enzyme was sufficient for thiamine but not for purine synthesis. The respective pseudorevertants were found to be informational suppressors. In addition, it was shown that accumulation of the purine intermediate aminoimidazole carboxamide ribotide inhibits thiamine synthesis, specifically affecting the conversion of aminoimidazole ribotide to hydroxymethyl pyrimidine.
Figures
References
-
- Altschul, S. F., W. Gish, W. Miller, E. W. Meyers, and D. J. Lipman. 1990. Basic local alignment search tool. J. Mol. Biol. 215:403-410. - PubMed
-
- Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman, J. A. Smith, and K. Struhl. 1994. Current protocols in molecular biology. John Wiley & Sons, New York, N.Y.
-
- Barik, S. 1995. Site-directed mutagenesis by double polymerase chain reaction. Mol. Biotechnol. 3:1-7. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
