

Phenotypes of lexA mutations in Salmonella enterica: evidence for a lethal lexA null phenotype due to the Fels-2 prophage
- PMID: 12399494
- PMCID: PMC151935
- DOI: 10.1128/JB.184.22.6235-6249.2002
Phenotypes of lexA mutations in Salmonella enterica: evidence for a lethal lexA null phenotype due to the Fels-2 prophage
Abstract
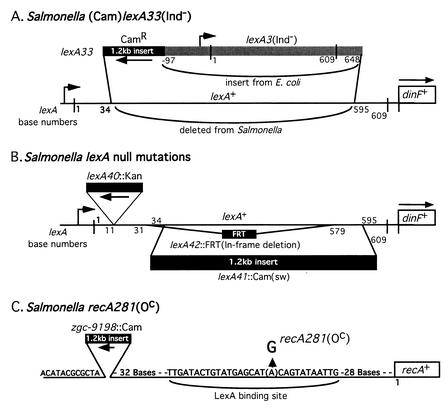
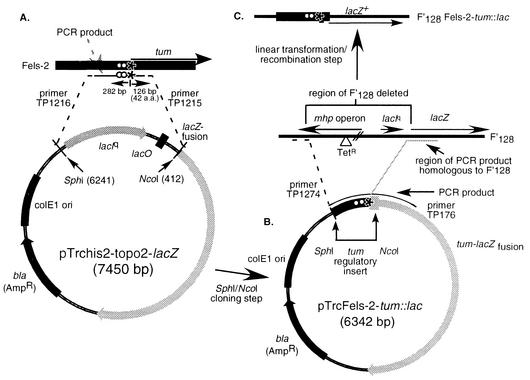
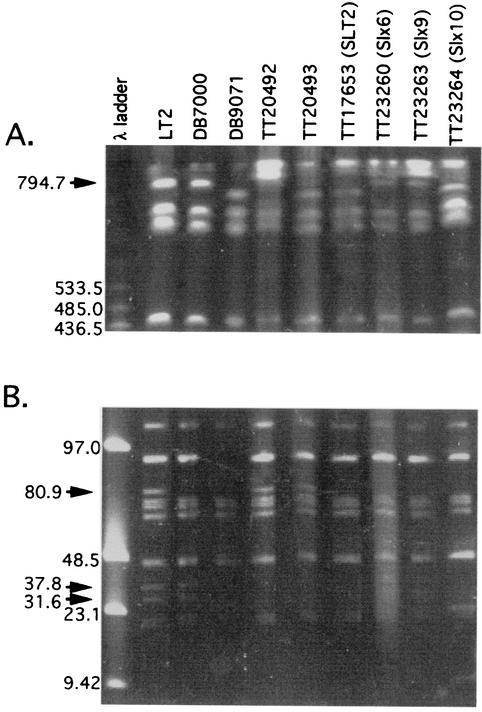
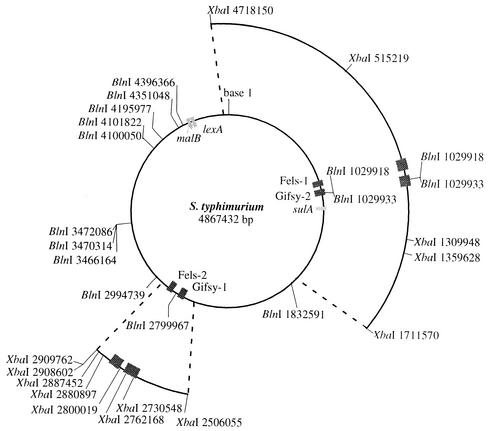
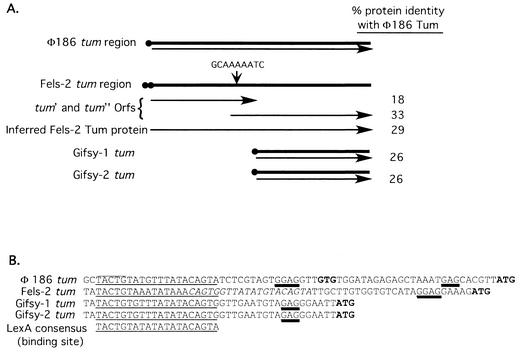
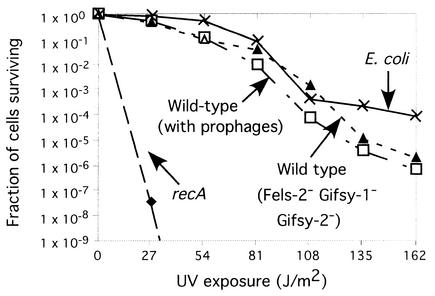
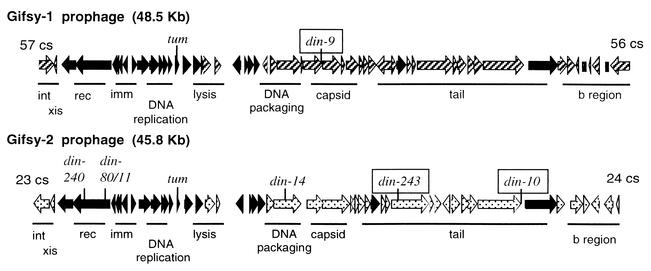
The LexA protein of Escherichia coli represses the damage-inducible SOS regulon, which includes genes for repair of DNA. Surprisingly, lexA null mutations in Salmonella enterica are lethal even with a sulA mutation, which corrects lexA lethality in E. coli. Nine suppressors of lethality isolated in a sulA mutant of S. enterica had lost the Fels-2 prophage, and seven of these (which grew better) had also lost the Gifsy-1 and Gifsy-2 prophages. All three phage genomes included a homologue of the tum gene of coliphage 186, which encodes a LexA-repressed cI antirepressor. The tum homologue of Fels-2 was responsible for lexA lethality and had a LexA-repressed promoter. This basis of lexA lethality was unexpected because the four prophages of S. enterica LT2 are not strongly UV inducible and do not sensitize strains to UV killing. In S. enterica, lexA(Ind(-)) mutants have the same phenotypes as their E. coli counterparts. Although lexA null mutants express their error-prone DinB polymerase constitutively, they are not mutators in either S. enterica or E. coli.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
