

Control of outer vestibule dynamics and current magnitude in the Kv2.1 potassium channel
- PMID: 12407083
- PMCID: PMC2229558
- DOI: 10.1085/jgp.20028639
Control of outer vestibule dynamics and current magnitude in the Kv2.1 potassium channel
Abstract
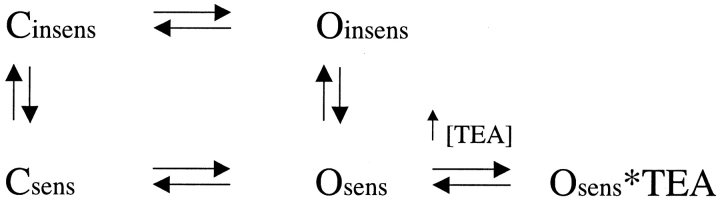
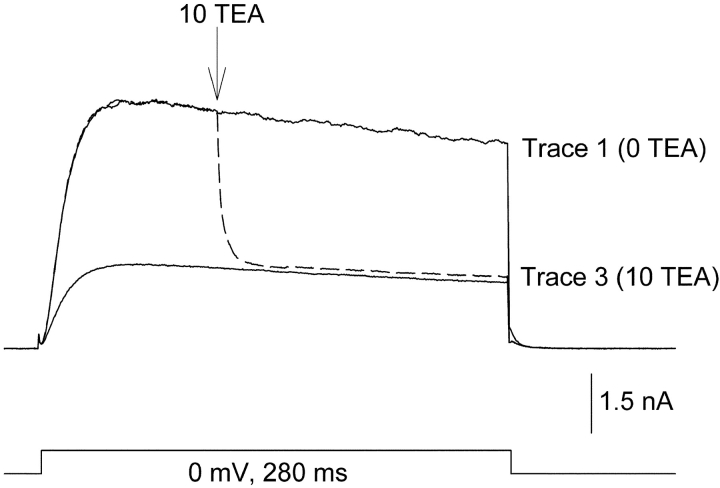
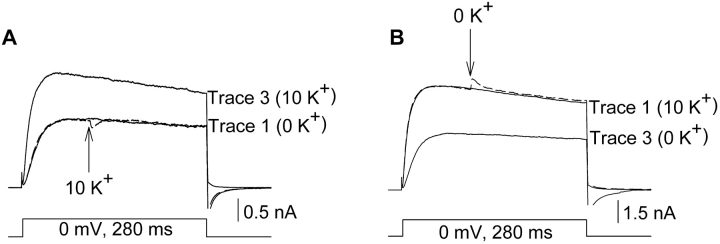
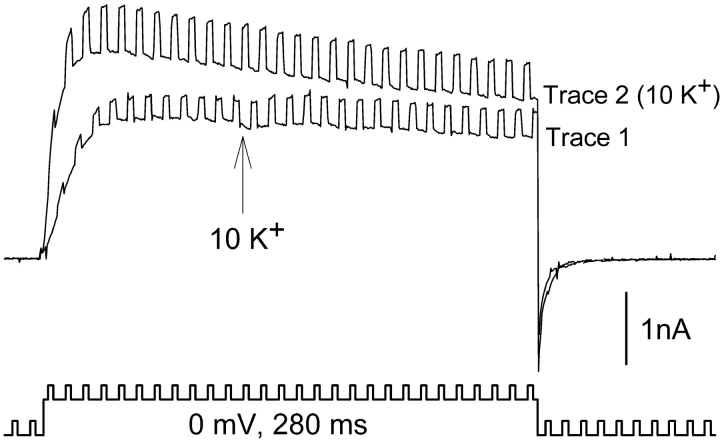
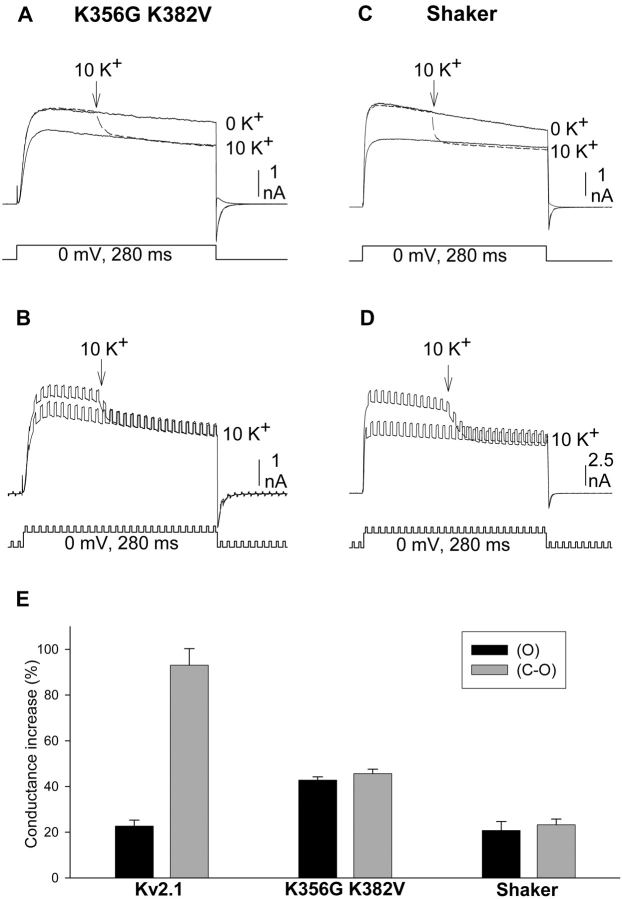
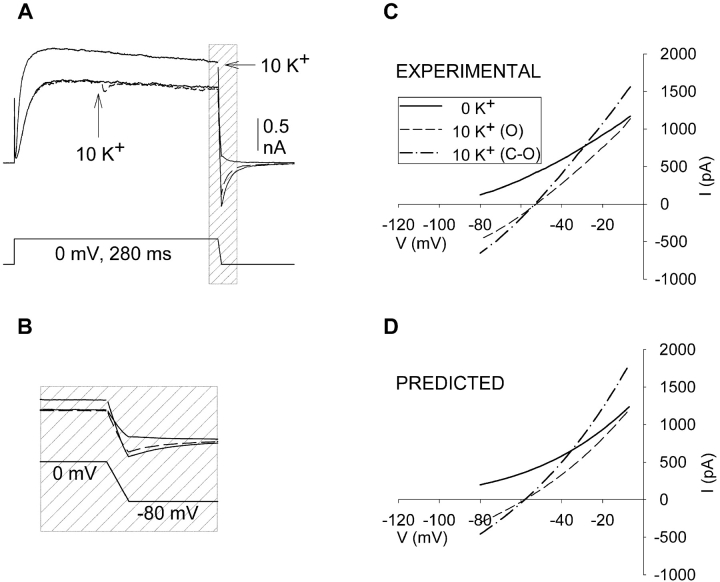
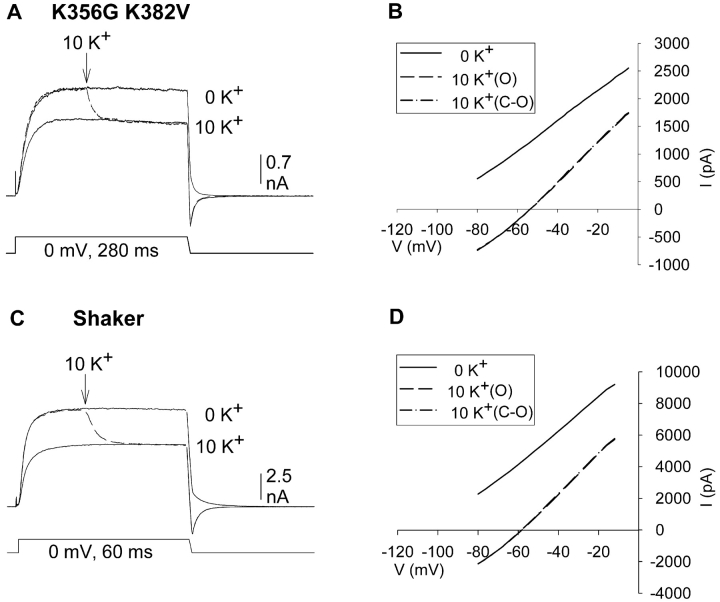
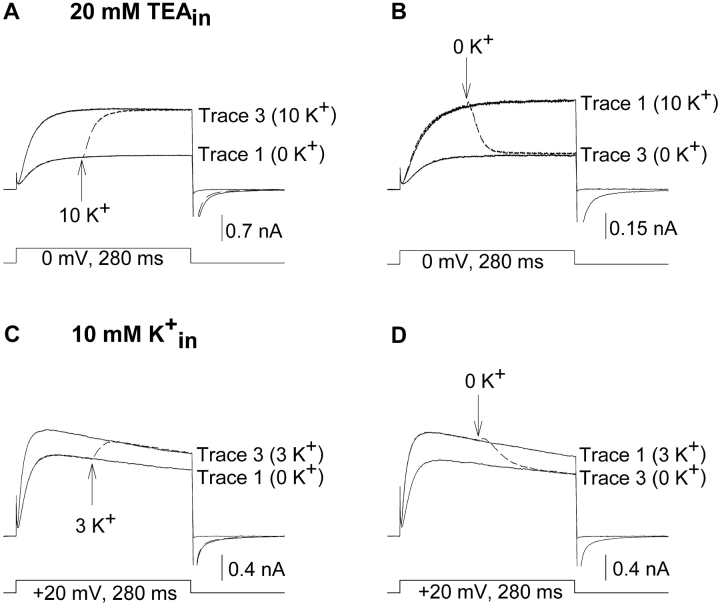
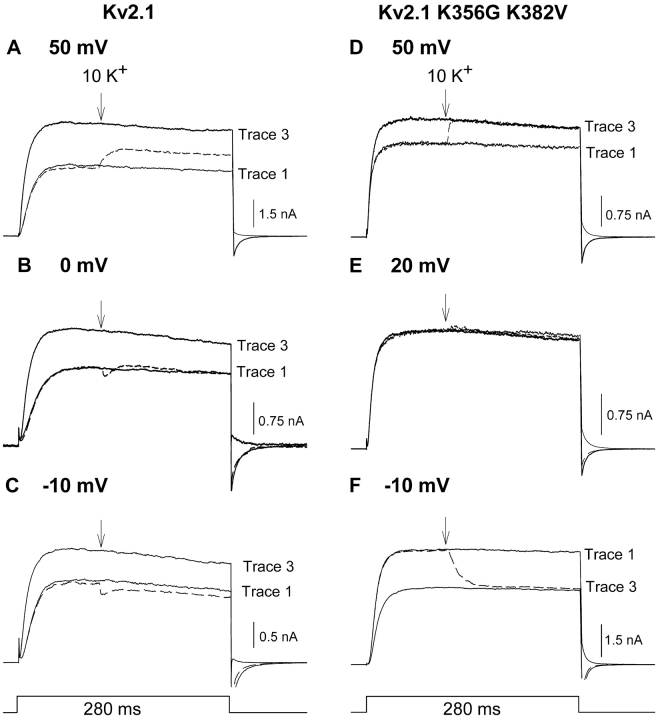
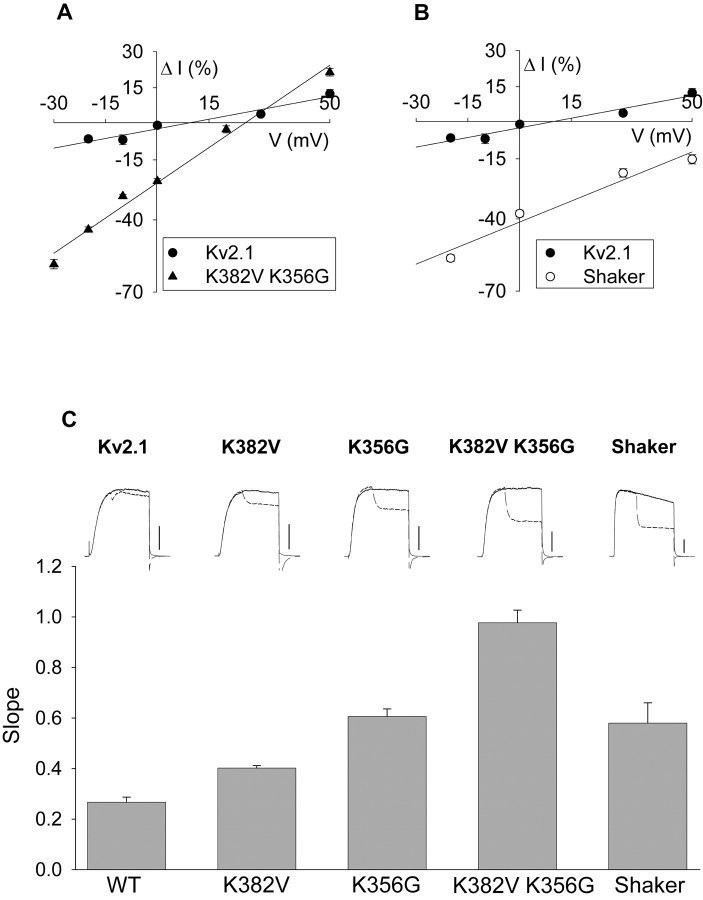
In Kv2.1 potassium channels, changes in external [K+] modulate current magnitude as a result of a K+-dependent interconversion between two outer vestibule conformations. Previous evidence indicated that outer vestibule conformation (and thus current magnitude) is regulated by the occupancy of a selectivity filter binding site by K+. In this paper, we used the change in current magnitude as an assay to study how the interconversion between outer vestibule conformations is controlled. With 100 mM internal K+, rapid elevation of external [K+] from 0 to 10 mM while channels were activated produced no change in current magnitude (outer vestibule conformation did not change). When channels were subsequently closed and reopened in the presence of elevated [K+], current magnitude was increased (outer vestibule conformation had changed). When channels were activated in the presence of low internal [K+], or when K+ flow into conducting channels was transiently interrupted by an internal channel blocker, increasing external [K+] during activation did increase current magnitude (channel conformation did change). These data indicate that, when channels are in the activated state under physiological conditions, the outer vestibule conformation remains fixed despite changes in external [K+]. In contrast, when channel occupancy is lowered, (by channel closing, an internal blocker or low internal [K+]), the outer vestibule can interconvert between the two conformations. We discuss evidence that the ability of the outer vestibule conformation to change is regulated by the occupancy of a nonselectivity filter site by K+. Independent of the outer vestibule-based potentiation mechanism, Kv2.1 was remarkably insensitive to K+-dependent processes that influence current magnitude (current magnitude changed by <7% at membrane potentials between -20 and 30 mV). Replacement of two outer vestibule lysines in Kv2.1 by smaller neutral amino acids made current magnitude dramatically more sensitive to the reduction in K+ driving force (current magnitude changed by as much as 40%). When combined, these outer vestibule properties (fixed conformation during activation and the presence of lysines) all but prevent variation in Kv2.1 current magnitude when [K+] changes during activation. Moreover, the insensitivity of Kv2.1 current magnitude to changes in K+ driving force promotes a more uniform modulation of current over a wide range of membrane potentials by the K+-dependent regulation of outer vestibule conformation.
Figures
References
-
- Baukrowitz, T., and G. Yellen. 1996. Use-dependent blockers and exit rate of the last ion from the multi-ion pore of a K+ channel. Science. 271:653–656. - PubMed
-
- Doyle, D.A., J.M. Cabral, R.A. Pfuetzner, A. Kuo, J.M. Gulbis, S.L. Cohen, B.T. Chait, and R. MacKinnon. 1998. The structure of the potassium channel: molecular basis of K+ conduction and selectivity. Science. 280:69–77. - PubMed
