

The cytoplasmic domain of the LDL receptor-related protein regulates multiple steps in APP processing
- PMID: 12411487
- PMCID: PMC131065
- DOI: 10.1093/emboj/cdf568
The cytoplasmic domain of the LDL receptor-related protein regulates multiple steps in APP processing
Abstract
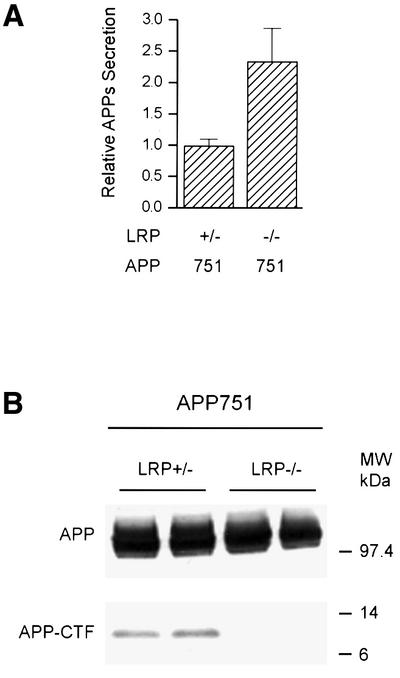
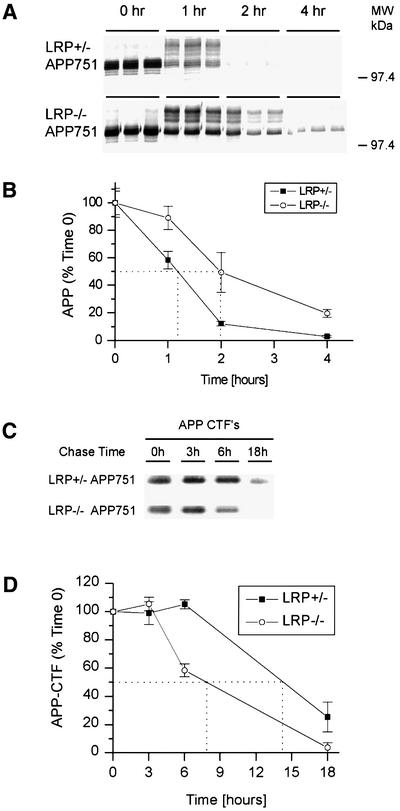
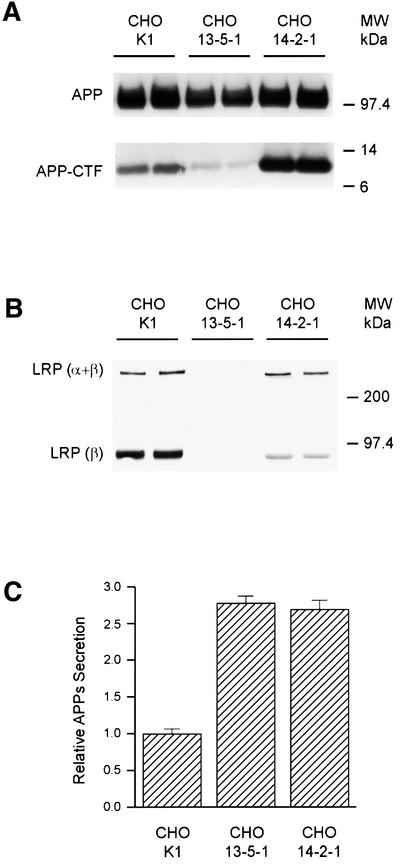
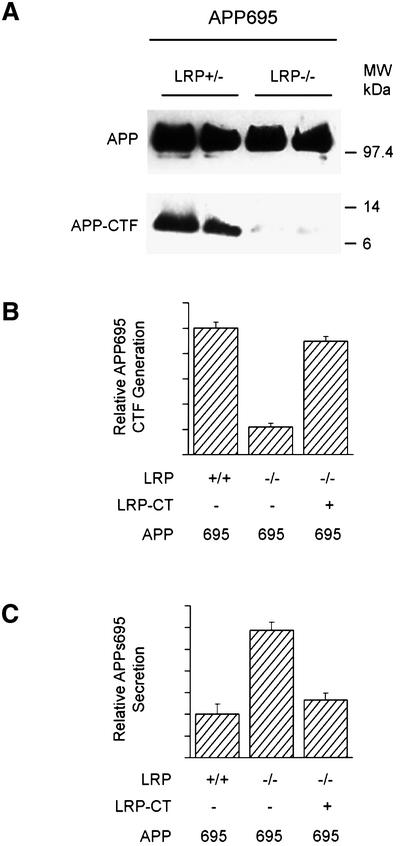
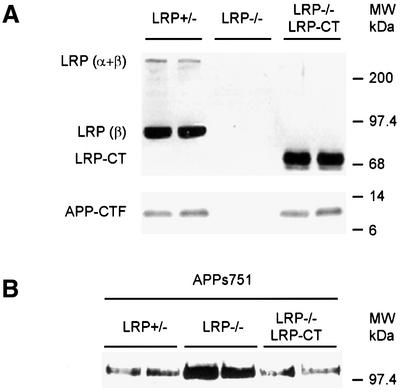
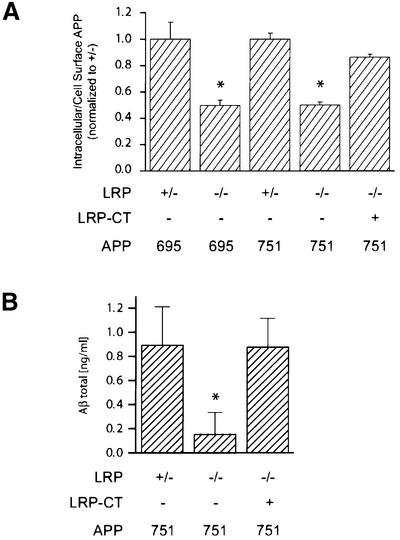
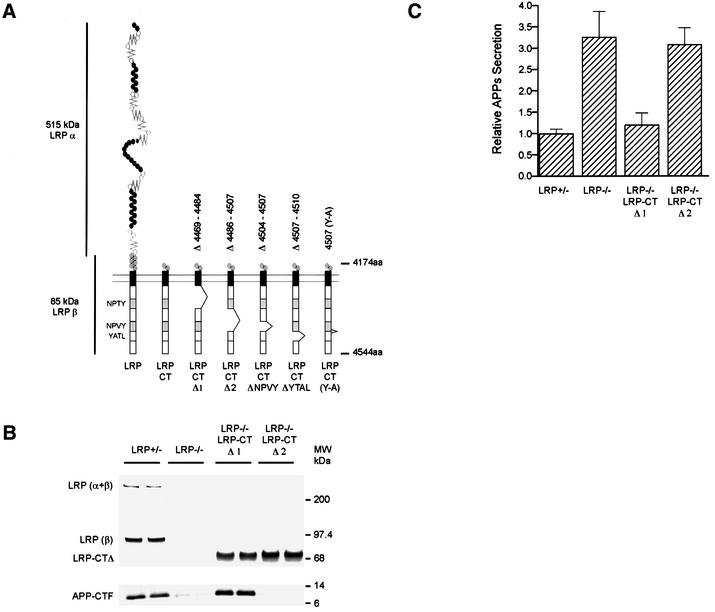
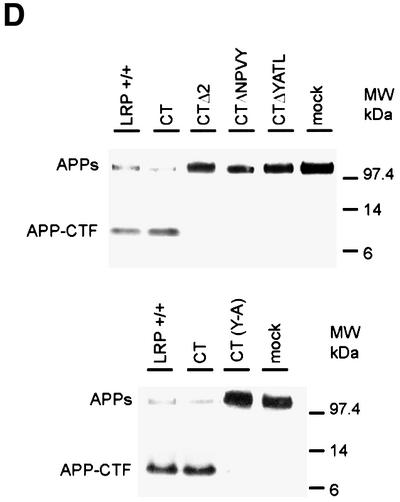
The low-density lipoprotein receptor-related protein (LRP) has recently been implicated in numerous intracellular signaling functions, as well as in Alzheimer's disease pathogenesis. Studies have shown that the beta-amyloid precursor protein (APP) interacts with LRP and that this association may impact the production of amyloid beta-protein (Abeta). In this report, we provide evidence that LRP regulates trafficking of intracellular proteins independently of its lipoprotein receptor functions. We show that in the absence of LRP, Abeta production, APP secretion, APP internalization, turnover of full-length APP and stability of APP C-terminal fragments are affected. Importantly, these changes are not APP isoform dependent. Using deletion constructs, the critical region in LRP that modulates APP processing was mapped to a seven peptide domain around the second NPXY domain (residues 4504-4510). Therefore, we propose a model by which LRP functionally modulates APP processing, including those steps critical for Abeta production, through interactions of the cytosolic domains.
Figures
References
-
- Boucher P., Liu,P., Gotthardt,M. Hiesberger,T., Anderson,R.G. and Herz,J. (2002) Platelet-derived growth factor mediates tyrosine phosphorylation of the cytoplasmic domain of the low density lipoprotein receptor-related protein in caveolae. J. Biol. Chem., 277, 15507–15513. - PubMed
-
- Bu G. and Rennke,S. (1996) Receptor-associated protein is a folding chaperone for low density lipoprotein receptor-related protein. J. Biol. Chem., 271, 22218–22224. - PubMed
-
- Cao X. and Sudhof,T.C. (2001) A transcriptively active complex of APP with Fe65 and histone acetyltransferase Tip60. Science, 293, 115–120. - PubMed
-
- Cook D.G., Forman,M.S., Sung,J.C., Leight,S., Kolson,D.L., Iwatsubo,T., Lee,V.M. and Doms,R.W. (1997) Alzheimer’s A β(1–42) is generated in the endoplasmic reticulum/intermediate compartment of NT2N cells. Nat. Med., 3, 1021–1023. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
