

Recombinant respiratory syncytial virus with the G and F genes shifted to the promoter-proximal positions
- PMID: 12414935
- PMCID: PMC136893
- DOI: 10.1128/jvi.76.23.11931-11942.2002
Recombinant respiratory syncytial virus with the G and F genes shifted to the promoter-proximal positions
Abstract
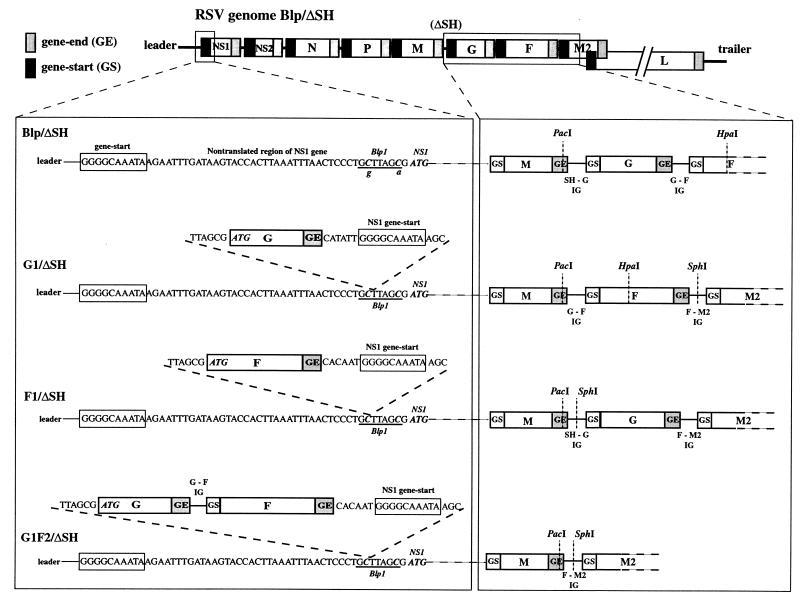
The genome of human respiratory syncytial virus (RSV) encodes 10 mRNAs and 11 proteins in the order 3'-NS1-NS2-N-P-M-SH-G-F-M2-1/M2-2-L-5'. The G and F glycoproteins are the major RSV neutralization and protective antigens. It seems likely that a high level of expression of G and F would be desirable for a live RSV vaccine. For mononegaviruses, the gene order is a major factor controlling the level of mRNA and protein expression due to the polar gradient of sequential transcription. In order to increase the expression of G and F, recombinant RSVs based on strain A2 were constructed in which the G or F gene was shifted from the sixth or seventh position (in a genome lacking the SH gene), respectively, to the first position (rRSV-G1/DeltaSH and rRSV-F1/DeltaSH, respectively). Another virus was made in which G and F were shifted together to the first and second positions, respectively (rRSV-G1F2/DeltaSH). Shifting one or two genes to the promoter-proximal position resulted in increased mRNA and protein expression of the shifted genes, with G and F expression increased up to 2.4-and 7.8-fold, respectively, at the mRNA level and approximately 2.5-fold at the protein level, compared to the parental virus. Interestingly, the transcription of downstream genes was not greatly affected even though shifting G or F, or G and F together, had the consequence of moving the block of genes NS1-NS2-N-P-M-(G) one or two positions further from the promoter. The efficiency of replication of the gene shift viruses in vitro was increased up to 10-fold. However, their efficiency of replication in the lower respiratory tracts of mice was statistically indistinguishable from that of the parental virus. In the upper respiratory tract, replication was slightly reduced on some days for viruses in which G was in the first position. The magnitude of the G-specific antibody response to the gene shift viruses was similar to that to the parental virus, whereas the F-specific response was increased up to fourfold, although this was not reflected in an increase of the neutralizing activity. Thus, shifting the G and F genes to the promoter-proximal position increased virus replication in vitro, had little effect on replication in the mouse, and increased the antigen-specific immunogenicity of the virus beyond that of parental RSV.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
