

Generation of hepatitis C virus-like particles by use of a recombinant vesicular stomatitis virus vector
- PMID: 12414973
- PMCID: PMC136870
- DOI: 10.1128/jvi.76.23.12325-12334.2002
Generation of hepatitis C virus-like particles by use of a recombinant vesicular stomatitis virus vector
Abstract
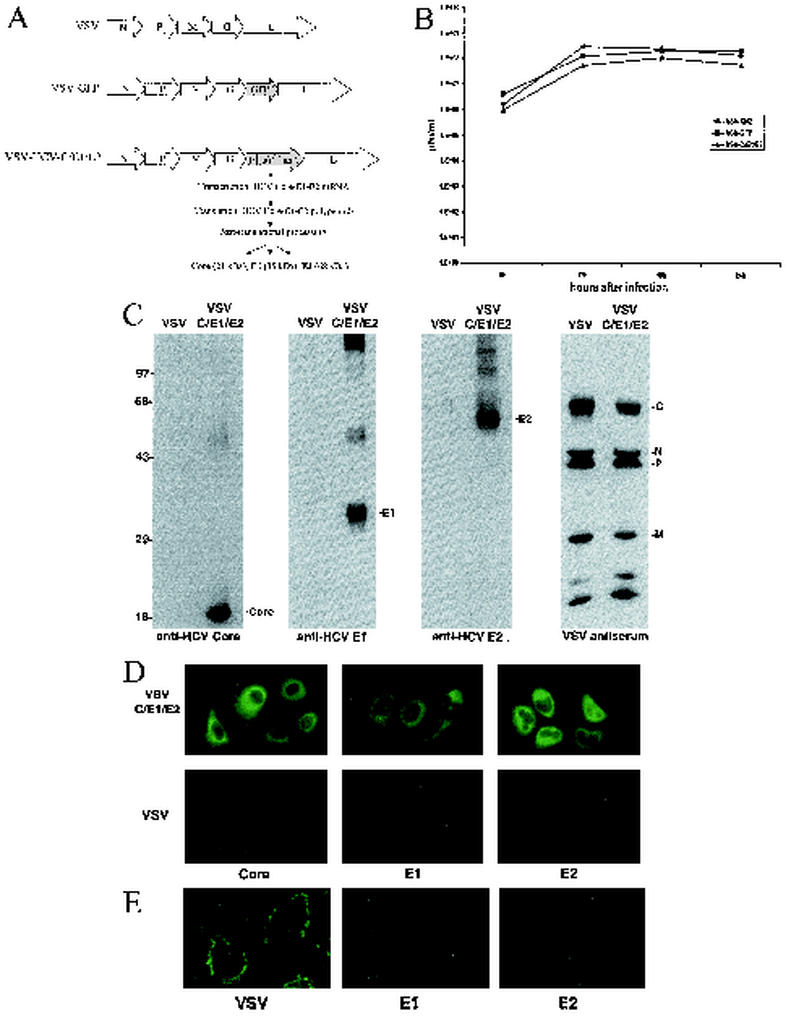
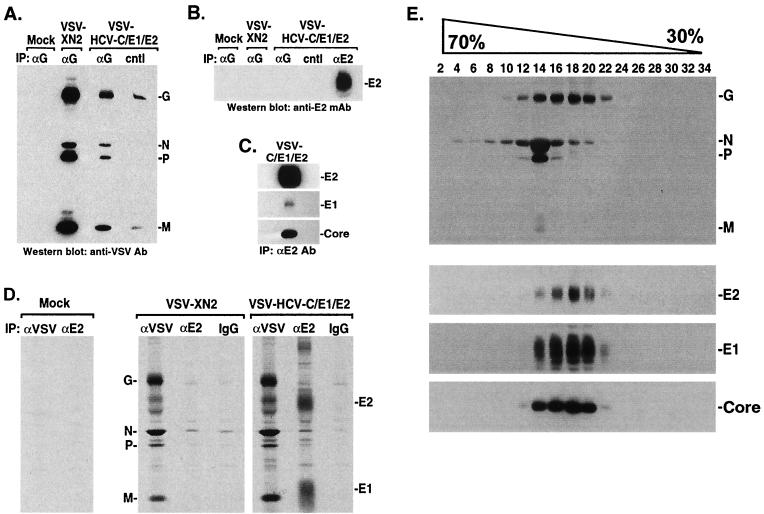
Hepatitis C virus (HCV), a major etiologic agent of hepatocellular carcinoma, presently infects approximately 400 million people worldwide, making the development of protective measures against HCV infection a key objective. Here we have generated a recombinant vesicular stomatitis virus (VSV), which expresses the HCV structural proteins, by inserting the contiguous Core, E1, and E2 coding region of HCV into the VSV genome. Recombinant VSV expressing HCV Core, E1, and E2 (VSV-HCV-C/E1/E2) grew to high titers in vitro and efficiently expressed the incorporated HCV gene product, which became fully processed into the individual HCV structural proteins. Biochemical and biophysical analysis indicated that the HCV Core, E1, and E2 proteins assembled to form HCV-like particles (HCV-LPs) possessing properties similar to the ultrastructural properties of HCV virions. Mice immunized with VSV-HCV-C/E1/E2 generated cell-mediated immune responses to all of the HCV structural proteins, and humoral responses, particularly to E2, were also readily evident. Our data collectively indicate that engineered VSVs expressing HCV Core, E1, and E2 and/or HCV-LPs represent useful tools in vaccine and immunotherapeutic strategies designed to address HCV infection.
Figures
Comment in
-
Endogenous virus and hepatitis C virus-like particle budding in BHK-21 cells.J Virol. 2003 Mar;77(6):3888-9; author reply 3889. doi: 10.1128/jvi.77.6.3888-3889.2003. J Virol. 2003. PMID: 12610167 Free PMC article. No abstract available.
References
-
- Adkins, B., Y. Bu, and P. Guevara. 2001. The generation of Th memory in neonates versus adults: prolonged primary Th2 effector function and impaired development of Th1 memory effector function in murine neonates. J. Immunol. 166:918-925. - PubMed
-
- Aizaki, H., Y. Aoki, T. Harada, K. Ishii, T. Suzuki, S. Nagamori, G. Toda, Y. Matsuura, and T. Miyamura. 1998. Full-length complementary DNA of hepatitis C virus genome from an infectious blood sample. Hepatology 27:621-627. - PubMed
-
- Arichi, T., T. Saito, M. E. Major, I. M. Belyakov, M. Shirai, V. H. Engelhard, S. M. Feinstone, and J. A. Berzofsky. 2000. Prophylactic DNA vaccine for hepatitis C virus (HCV) infection: HCV-specific cytotoxic T lymphocyte induction and protection from HCV-recombinant vaccinia infection in an HLA-A2.1 transgenic mouse model. Proc. Natl. Acad. Sci. USA 97:297-302. - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
