

doi: 10.1128/jvi.76.23.12335-12343.2002.
Binding of Norwalk virus-like particles to ABH histo-blood group antigens is blocked by antisera from infected human volunteers or experimentally vaccinated mice
Affiliations
- PMID: 12414974
- PMCID: PMC136916
- DOI: 10.1128/jvi.76.23.12335-12343.2002
Item in Clipboard
Binding of Norwalk virus-like particles to ABH histo-blood group antigens is blocked by antisera from infected human volunteers or experimentally vaccinated mice
J Virol.
2002 Dec.
Abstract
Attachment of Norwalk (NV), Snow Mountain (SMV), and Hawaii (HV) virus-like particles (VLPs) to specific ABH histo-blood group antigens was investigated by using human saliva and synthetic biotinylated carbohydrates. The three distinct Norwalk-like viruses (NLVs) have various capacities for binding ABH histo-blood group antigens, suggesting that different mechanisms for NLV attachment likely exist. Importantly, antisera from NV-infected human volunteers, as well as from mice inoculated with packaged Venezuelan equine encephalitis virus replicons expressing NV VLPs, blocked the ability of NV VLPs to bind synthetic H type 1, Le(b), and H type 3, suggesting a potential mechanism for antibody-mediated neutralization of NV.
Figures
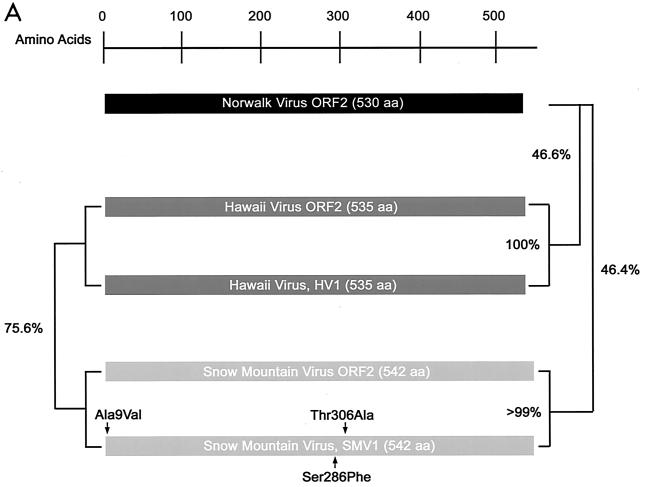
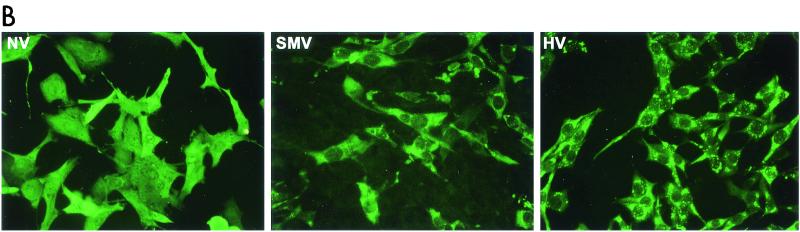
Expression and self-assembly of NLV capsid proteins. HV and SMV genomic RNA was isolated from the stools of SMV- or HV-infected human volunteers. The capsid genes were isolated by reverse transcription-PCR and were subcloned into the pVR21 VEE replicon vector as previously described for NV1 (3) with specific primer pairs 5′-AGTCTAGTCCGCCAAGATGAAGATGGCGTCGAATGAC-3′ and 5′-NNNTTAATTAATTATTGCACTCTTCTGCGCC-3′ (HV-5′ and HV-3′, respectively) for HV or HV-5′ and 5′-NNNNNNNGGCGCGCCTTACTGAACCCTTCTACGC-3′ (SMV-3′) for SMV. (A) Amino acid (aa) alignment of NV, HV, and SMV ORF2 regions. NV is a genogroup I isolate, whereas SMV and HV are genogroup II NLVs from distinct genogroup clusters (GII.2 and GII.1, respectively). Percentage of amino acid identities are shown, and arrows indicate amino acid variations from published sequences (12-14). The NV ORF2 capsid clone NV1 has been previously described and is identical to the published NV ORF2 amino acid sequence (9, 12). (B) BHK cells were infected with packaged VRPs encoding NV, SMV, or HV capsid proteins. Expression of NLV capsid proteins was determined by IFA with human antiserum directed to either NV, SMV, or HV. After determination of the VRP titers by IFA as described previously (3, 9), BHK cells were infected with VRPs encoding either NV, SMV, or HV capsid proteins at a multiplicity of infection of 2. At 36 h postinfection BHK cells were lysed by freeze-thaw and the extracts were purified through sucrose gradients and were analyzed by negative-stain electron microscopic analysis (C). Scale bar, 100 nm.
Expression and self-assembly of NLV capsid proteins. HV and SMV genomic RNA was isolated from the stools of SMV- or HV-infected human volunteers. The capsid genes were isolated by reverse transcription-PCR and were subcloned into the pVR21 VEE replicon vector as previously described for NV1 (3) with specific primer pairs 5′-AGTCTAGTCCGCCAAGATGAAGATGGCGTCGAATGAC-3′ and 5′-NNNTTAATTAATTATTGCACTCTTCTGCGCC-3′ (HV-5′ and HV-3′, respectively) for HV or HV-5′ and 5′-NNNNNNNGGCGCGCCTTACTGAACCCTTCTACGC-3′ (SMV-3′) for SMV. (A) Amino acid (aa) alignment of NV, HV, and SMV ORF2 regions. NV is a genogroup I isolate, whereas SMV and HV are genogroup II NLVs from distinct genogroup clusters (GII.2 and GII.1, respectively). Percentage of amino acid identities are shown, and arrows indicate amino acid variations from published sequences (12-14). The NV ORF2 capsid clone NV1 has been previously described and is identical to the published NV ORF2 amino acid sequence (9, 12). (B) BHK cells were infected with packaged VRPs encoding NV, SMV, or HV capsid proteins. Expression of NLV capsid proteins was determined by IFA with human antiserum directed to either NV, SMV, or HV. After determination of the VRP titers by IFA as described previously (3, 9), BHK cells were infected with VRPs encoding either NV, SMV, or HV capsid proteins at a multiplicity of infection of 2. At 36 h postinfection BHK cells were lysed by freeze-thaw and the extracts were purified through sucrose gradients and were analyzed by negative-stain electron microscopic analysis (C). Scale bar, 100 nm.
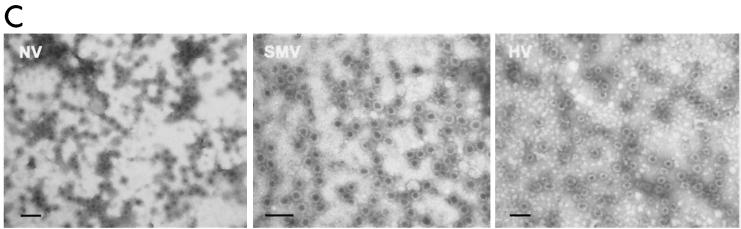
Expression and self-assembly of NLV capsid proteins. HV and SMV genomic RNA was isolated from the stools of SMV- or HV-infected human volunteers. The capsid genes were isolated by reverse transcription-PCR and were subcloned into the pVR21 VEE replicon vector as previously described for NV1 (3) with specific primer pairs 5′-AGTCTAGTCCGCCAAGATGAAGATGGCGTCGAATGAC-3′ and 5′-NNNTTAATTAATTATTGCACTCTTCTGCGCC-3′ (HV-5′ and HV-3′, respectively) for HV or HV-5′ and 5′-NNNNNNNGGCGCGCCTTACTGAACCCTTCTACGC-3′ (SMV-3′) for SMV. (A) Amino acid (aa) alignment of NV, HV, and SMV ORF2 regions. NV is a genogroup I isolate, whereas SMV and HV are genogroup II NLVs from distinct genogroup clusters (GII.2 and GII.1, respectively). Percentage of amino acid identities are shown, and arrows indicate amino acid variations from published sequences (12-14). The NV ORF2 capsid clone NV1 has been previously described and is identical to the published NV ORF2 amino acid sequence (9, 12). (B) BHK cells were infected with packaged VRPs encoding NV, SMV, or HV capsid proteins. Expression of NLV capsid proteins was determined by IFA with human antiserum directed to either NV, SMV, or HV. After determination of the VRP titers by IFA as described previously (3, 9), BHK cells were infected with VRPs encoding either NV, SMV, or HV capsid proteins at a multiplicity of infection of 2. At 36 h postinfection BHK cells were lysed by freeze-thaw and the extracts were purified through sucrose gradients and were analyzed by negative-stain electron microscopic analysis (C). Scale bar, 100 nm.
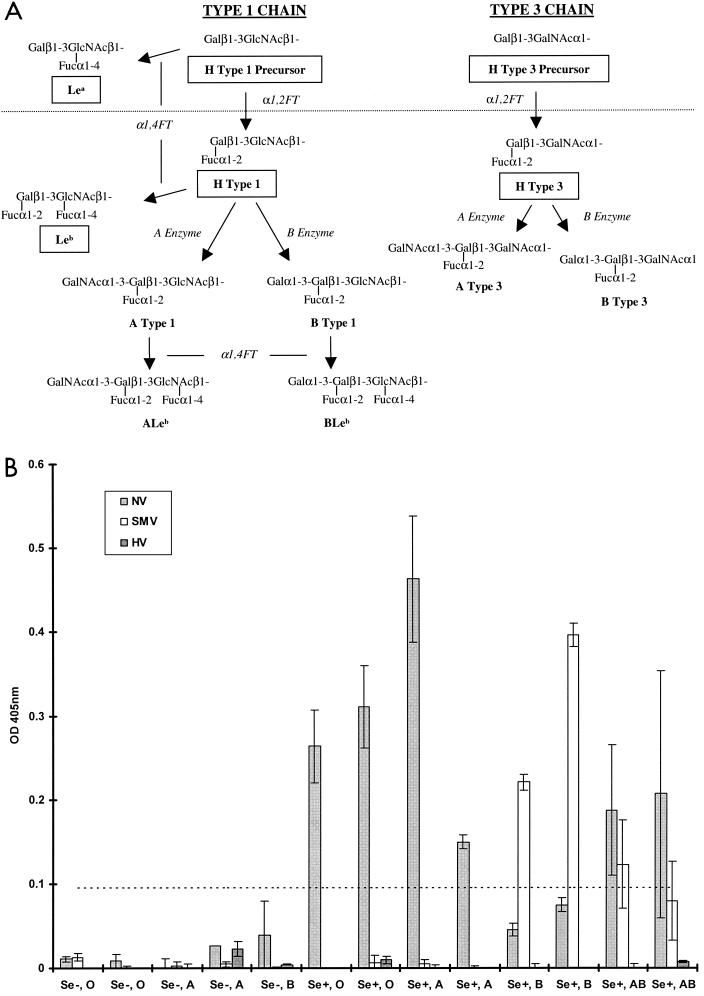
Binding of NLV VLPs to saliva components from individuals of different blood types and secretor status. (A) General schematic of the type 1 and 3 chain ABH histo-blood group antigen production pathways adapted from that described in reference . Synthetic carbohydrates used in this paper are boxed. Enzymatic activities are in italics (FT, fucosyltransferase). The type 1 and 3 chains are the primary histo-blood group carbohydrates present in saliva and on epithelial cell surfaces of the gastric mucosa, presumed to be the site of NLV attachment (16, 20). H antigens are the first carbohydrates synthesized from precursor disaccharides by the enzymatic activity of the α1,2 fucosyltransferase encoded by either the fut1 or fut2 gene. About 20% of the European population encodes a mutant form of the fut2 gene resulting in the nonsecretor phenotype. Nonsecretors do not express a functional FUT2 enzyme and will thus not produce carbohydrates downstream of the H precursors (dashed line) on gastric mucosal surfaces or in saliva. In secretors, the A or B enzyme expressed by blood type A or B individuals, respectively (or both in type AB individuals), can further modify the H antigens along the particular type chain. Individuals of blood type O do not produce A or B enzymes and therefore do not express A or B antigens or downstream carbohydrates on mucosal surfaces. The Lea and Leb antigens are synthesized by the FUT3 Lewis enzymes (α1-3/4 fucosyltransferases) from either the H type 1 precursor or H type 1, respectively. In saliva and in the gastric mucosa, Lewis-positive secretors will generally produce Leb and relatively less Lea, whereas nonsecretors will only produce Lea. (B) Binding of NLV VLPs to saliva from individuals of various secretor status and blood type. VLPs were added to saliva-coated microwells, and VLP binding was detected with appropriate human anti-NLV antisera, anti-IgG alkaline phosphatase conjugated secondary antibody, and pNPP substrate. The OD405 quantifies net VLP binding to saliva after background subtraction, and the dashed line represents an arbitrary negative cutoff value equivalent to twice the average background OD. Mean values from duplicate wells are shown, and error bars represent the standard deviation. The x axis displays the individuals' secretor phenotype and blood type. Note that saliva samples could only be obtained from one Se− blood type B and no Se− blood type AB individuals.
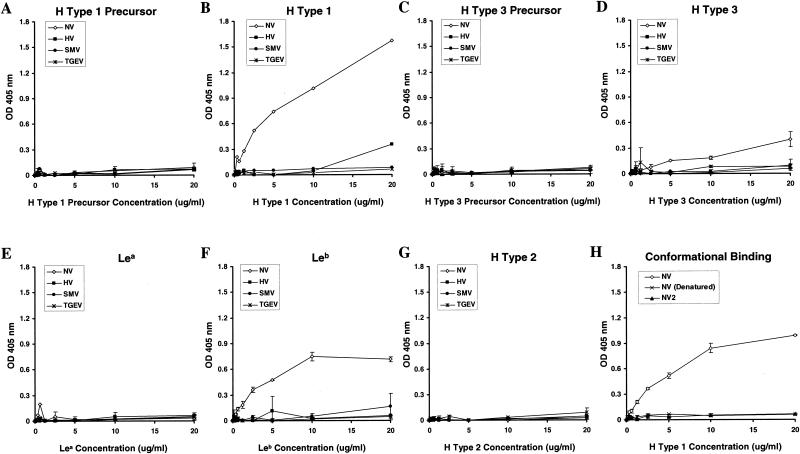
(A to G) Binding of synthetic ABH histo-blood group antigens with NLV VLPs. Purified NLV VLPs (100 μl at 2 μg/ml in TBS) were added to high-binding enzyme immunoassay plates (Costar, Corning, N.Y.). Sucrose-purified virions of the coronavirus transmissible gastroenteritis virus (TGEV) were used as an unrelated virus control. Plates were incubated for 4 h at ambient temperature and were blocked overnight at 4°C with 5% milk in phosphate-buffered saline (5% BLOTTO). Synthetic ABH and Lewis histo-blood group carbohydrates were purchased from Glycotech (Rockville, Md.). Each synthetic carbohydrate is biotinylated with a multivalent polyacrylamide linker between the biotin and carbohydrate moieties. The lyophylized carbohydrates were resuspended in sterile phosphate-buffered saline at 1 mg/ml, diluted to 20 μg/ml in 5% BLOTTO, and serially diluted twofold in the microtiter wells, yielding 100 μl/well. The plates were incubated for 3.5 to 4 h at 37°C followed by five washes with phosphate-buffered saline-Tween (0.05%). A 1:250 dilution (100 μl/well; in 5% BLOTTO) of a streptavidin-alkaline phosphatase conjugate solution (1 mg/ml; Sigma) was then added, and the plates were incubated at 37°C for 1 h. After a final wash step the plates were developed with 150 μl of pNPP substrate/well and the OD405 was determined. Wells incubated with blocking solution instead of synthetic carbohydrates were used as plate blanks. (H) Conformation-dependent binding of H type 1 with NV VLPs. High-binding microwells were coated with (2 μg/ml) NV VLPs (NV), heat denatured NV VLPs [NV (Denatured)], or sucrose-purified NV2 protein (NV2), which does not assemble into intact VLPs (9). Serially diluted, synthetic, biotinylated H type 1 was added to the microwells, and the ability of the carbohydrate to bind to the NV capsid proteins was determined. Average ODs for all panels are shown, and error bars represent the standard deviation of duplicate wells.
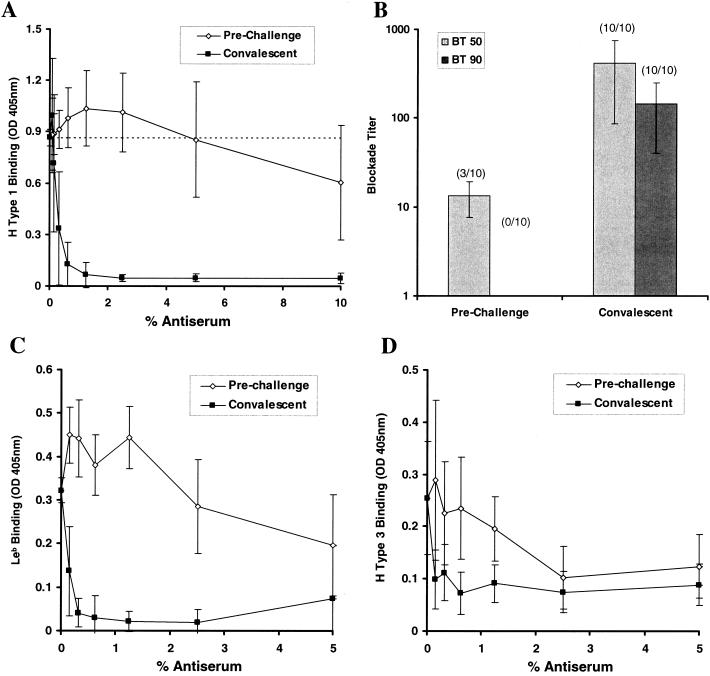
Human antisera blockade of NV VLP:ABH histo-blood group attachment. Prechallenge and convalescent-phase serum samples were obtained from 10 human volunteers infected with a live NV challenge inoculum. Sera were serially diluted and added to NV-coated microwells prior to addition of synthetic, biotinylated carbohydrates and completion of the binding assay. (A) The OD405 was determined to measure H type 1:NV VLP binding in the presence of various concentrations of the preincubated, prechallenge, and convalescent human antisera. Results indicate the mean OD405 from 10 human serum sample pairs, and error bars represent standard deviation. The dashed line represents 100% H type 1 binding (no preincubated serum). (B) The serum dilution (blockade titer) at which 50 and 90% H type 1 is blocked from binding to the NV VLPs (BT 50 and BT 90, respectively) was determined on the basis of OD values. Results indicate the mean blockade titer of the serum samples, with detectable BT 50 and BT 90 values at the concentrations tested, and the number of samples with detectable BT 50 or BT 90 titers is indicated by parentheses. Error bars indicate blockade titer standard deviation from only those samples with detectable BT 50 and BT 90 values. (C and D) Human antisera blockade of Leb:NV VLP and H type 3:NV VLP binding. Serum samples were preincubated with NV VLP-coated wells prior to addition of the biotinylated carbohydrates and completion of the binding assay. Results indicate mean OD values for 10 serum pairs, and error bars represent standard deviation.
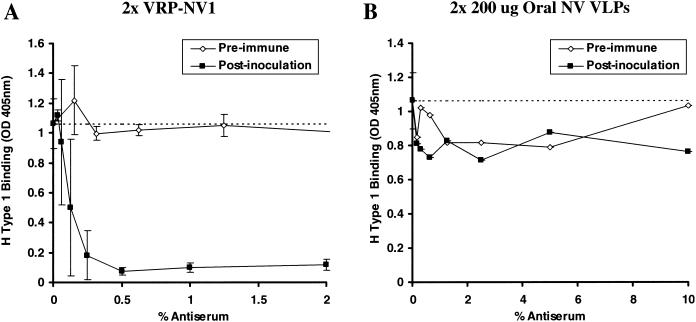
Mouse antisera blockade of H type 1:NV VLP binding. Groups of four mice were experimentally vaccinated with two doses of either subcutaneously inoculated VRP-NV1 or orally inoculated NV VLPs. VRPs were administered at 107 infectious units via footpad injection, and 200 μg of sucrose-purified NV VLPs was administered by oral gavage by using methods previously described (9). Baseline and day 35 (12 days postboost) antisera were serially diluted and preincubated in NV VLP coated microwells, followed by addition of synthetic, biotinylated H type 1. H type 1 binding was measured as described in the legend to Fig. 4 and in the text. Dashed lines represent 100% H type 1 binding (no preincubated serum). (A) Mean ODs from wells preincubated with the various serum concentrations from four mice inoculated with VRP-NV1. Error bars represent standard deviation. Note that data for preimmune sera from only three of the four VRP-NV1-vaccinated mice is shown because of insufficient sample volume from one mouse. (B) OD values from microwells preincubated with serially diluted sera from one mouse that responded to the orally inoculated NV VLPs. The three other mice did not respond to the experimental vaccination (9). Note the x-axis scale differences in panels A and B.
References
-
- Ball, J. M., D. Y. Graham, A. R. Opekun, M. A. Gilger, R. A. Guerrero, and M. K. Estes. 1999. Recombinant Norwalk virus-like particles given orally to volunteers: phase I study. Gastroenterology 117:40-48. - PubMed
-
- Edwards, M. J., and N. J. Dimmock. 2001. A haemagglutinin (HA1)-specific FAb neutralizes influenza A virus by inhibiting fusion activity. J. Gen. Virol. 82:1387-1395. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
