

RNA structure and packaging signals in the 5' leader region of the human immunodeficiency virus type 1 genome
- PMID: 12414982
- PMCID: PMC136901
- DOI: 10.1128/jvi.76.23.12381-12387.2002
RNA structure and packaging signals in the 5' leader region of the human immunodeficiency virus type 1 genome
Abstract
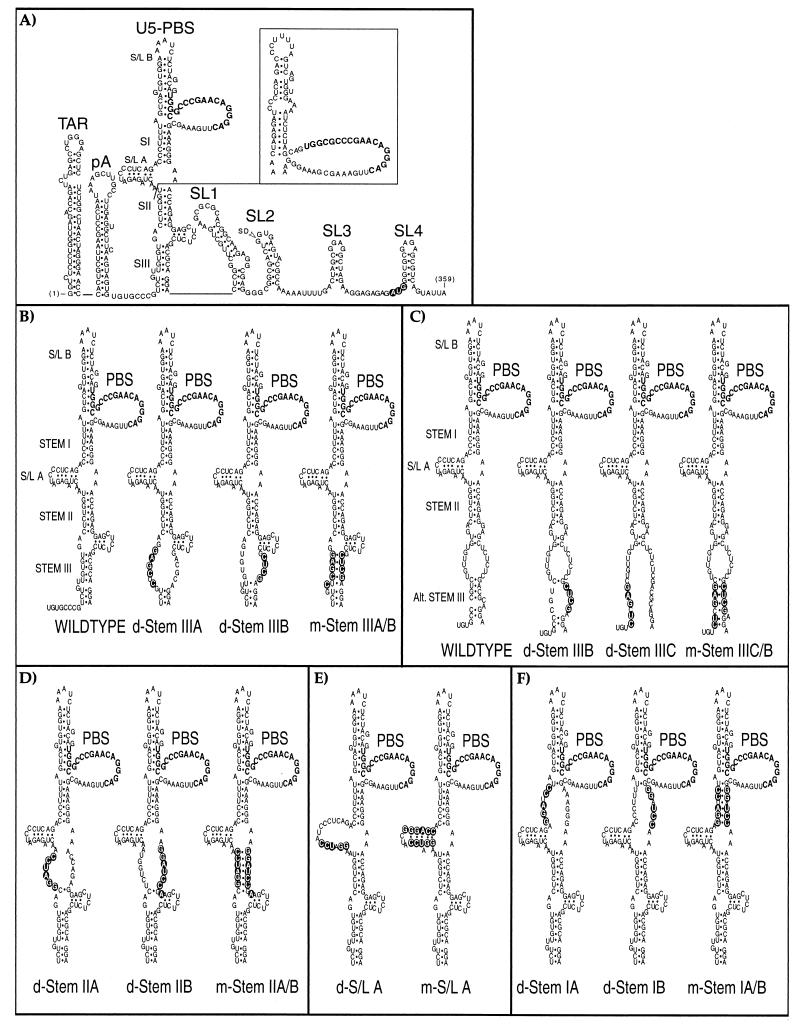
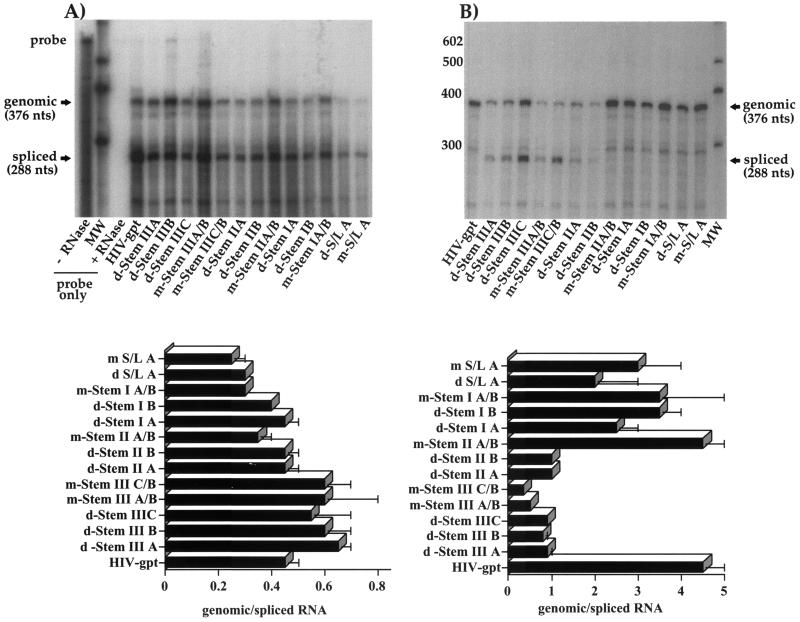
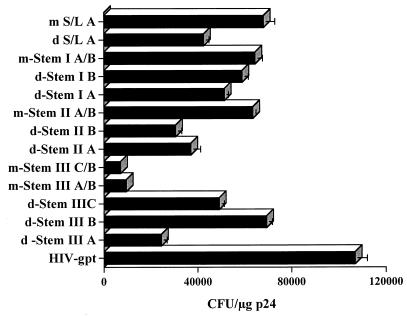
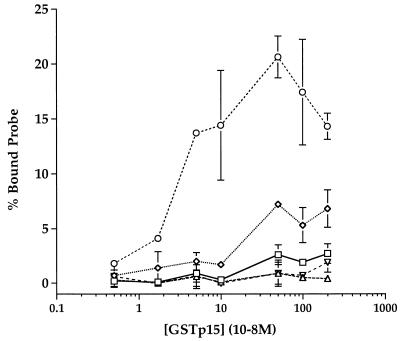
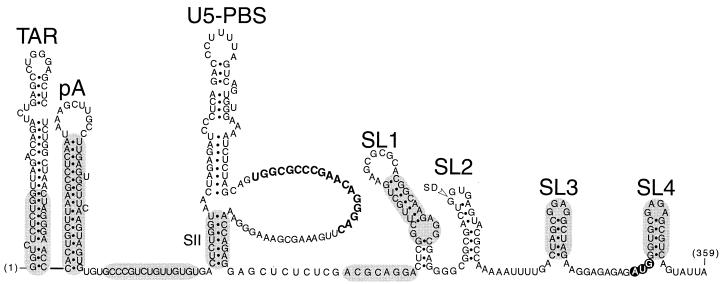
The leader region of the human immunodeficiency virus type 1 (HIV-1) genome has a highly folded structure, comprising at least two RNA stem-loops [the transactivation response (TAR) and poly(A) hairpins] near its 5' end and four others (SL1 to SL4) downstream. Each of these stem-loops contributes to the function of the HIV-1 packaging signal, which efficiently targets genomic RNA into nascent virions. The central 140-base region of the leader, which includes the U5 and primer binding site (PBS) sequences, is also believed to adopt a complex structure, but the nature of this structure and its possible role in RNA packaging have not been extensively explored. Here we report a mutational analysis identifying at least three separate loci within the U5-PBS region which, when mutated, impair both HIV-1 packaging specificity and infectivity in a single-round proviral assay. In common with those of all previously described packaging signals in the leader, the function of one of these loci appeared to depend on secondary structure rather than on sequence alone. By contrast, the activity of the other two loci did not correlate with any predicted conformations. Moreover, unlike SL1 to SL4, the TAR, poly(A), and U5-PBS hairpins were not bound with high affinity by the nucleocapsid portion of the HIV-1 Gag protein in vitro, implying that they contribute to packaging through a mechanism distinct from that of SL1 to SL4. Our findings confirm the existence and importance of secondary structure around the PBS and demonstrate that functional packaging signals are distributed across the entire HIV-1 leader.
Figures
References
-
- Beerens, N., F. Groot, and B. Berkhout. 2001. Initiation of HIV-1 reverse transcription is regulated by a primer activation signal. J. Biol. Chem. 276:31247-31256. - PubMed
-
- Berkhout, B. 1996. Structure and function of the human immunodeficiency virus leader RNA. Prog. Nucleic Acid Res. Mol. Biol. 54:1-34. - PubMed
-
- Berkowitz, R., J. Fisher, and S. P. Goff. 1996. RNA packaging. Curr. Top. Microbiol. Immunol. 214:177-218. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
