

VH1, a provascular cell-specific receptor kinase that influences leaf cell patterns in Arabidopsis
- PMID: 12417696
- PMCID: PMC152722
- DOI: 10.1105/tpc.005884
VH1, a provascular cell-specific receptor kinase that influences leaf cell patterns in Arabidopsis
Abstract
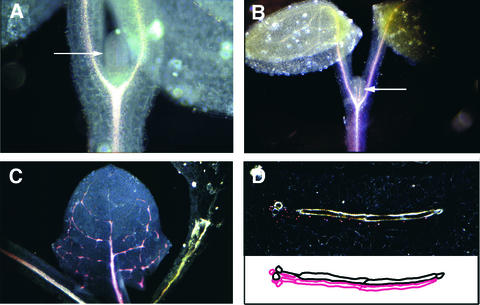
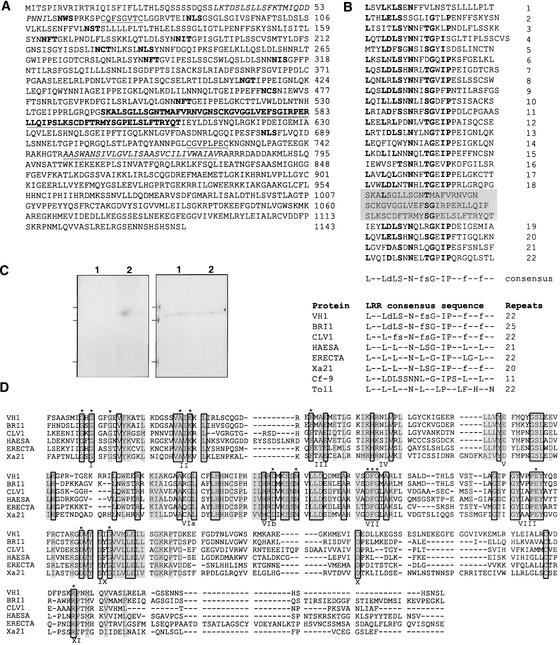
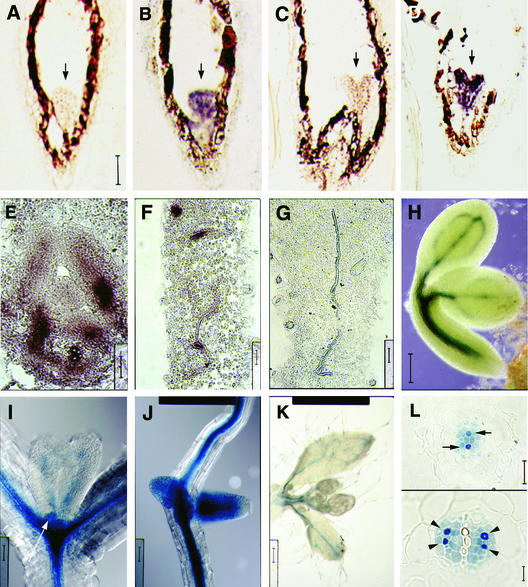
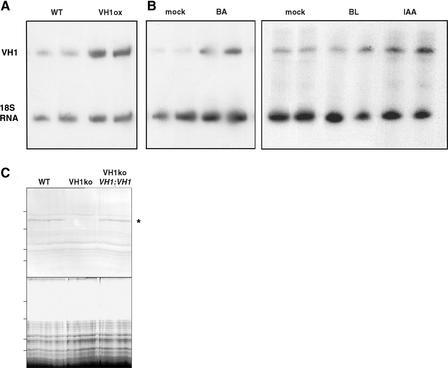
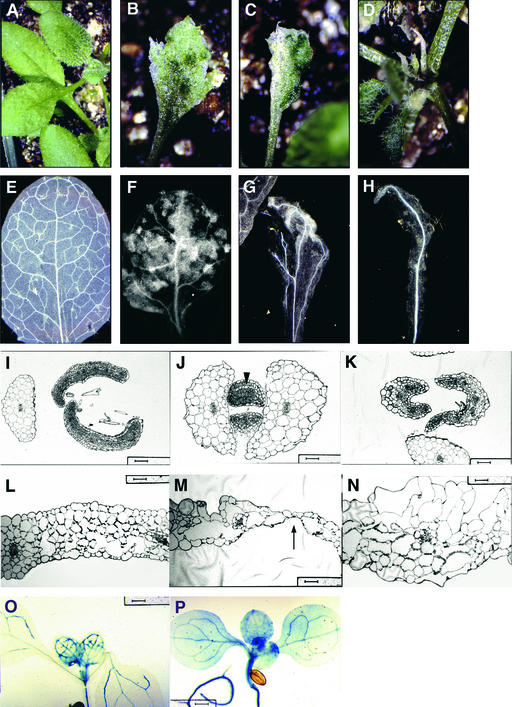
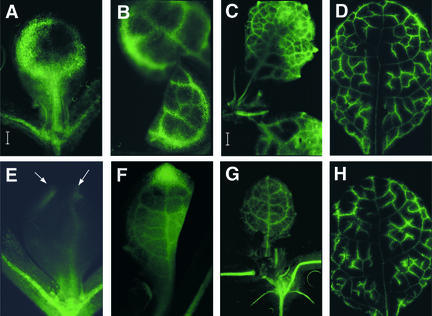
The formation of the venation pattern in leaves is ideal for examining signaling pathways that recognize and respond to spatial and temporal information, because the pattern is two-dimensional and heritable and the resulting veins influence the three-dimensional spatial organization of the surrounding differentiating leaf cell types. We identified a provascular/procambial cell-specific gene that encodes a Leu-rich repeat receptor kinase, which we named VASCULAR HIGHWAY1 (VH1). A change in the expression domain and level of VH1 marks the transition from an uncommitted provascular state to a committed procambial state in early vascular development. The coding sequence, expression pattern, and transgenic phenotypes together suggest that VH1 transduces extracellular spatial and temporal signals into downstream cell differentiation responses in provascular/procambial cells.
Figures

References
-
- Badenhuizen, N.P. (1969). The Biogenesis of Starch Granules in Higher Plants. (New York: Appleton-Century-Crofts).
-
- Baima, S., Nobili, F., Sessa, G., Luchetti, S., Ruberti, I., and Morelli, G. (1995). The expression of the Athb-8 homeobox gene is restricted to provascular cells in Arabidopsis thaliana. Development 121, 4171–4182. - PubMed
-
- Baker, B., Zambryski, P., Staskawicz, B., and Dinesh-Kumar, S.P. (1997). Signaling in plant-microbe interactions. Science 276, 726–733. - PubMed
-
- Berleth, T., and Jurgens, G. (1993). The role of the monopteros gene in organizing the basal body region of the Arabidopsis embryo. Development 118, 575–587.
-
- Berleth, T., Mattsson, J., and Hardtke, C.S. (2000). Vascular continuity and auxin signals. Trends Plant Sci. 5, 387–393. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
