

Engineered herpes simplex virus 1 is dependent on IL13Ralpha 2 receptor for cell entry and independent of glycoprotein D receptor interaction
- PMID: 12417744
- PMCID: PMC137554
- DOI: 10.1073/pnas.232588699
Engineered herpes simplex virus 1 is dependent on IL13Ralpha 2 receptor for cell entry and independent of glycoprotein D receptor interaction
Abstract
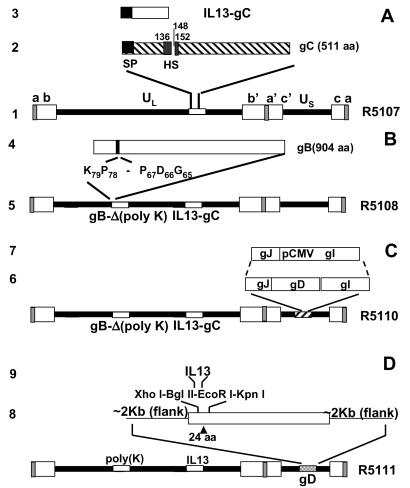
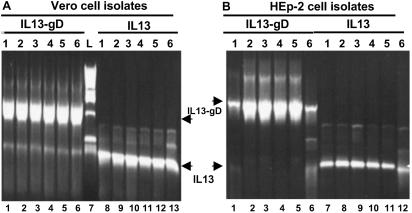
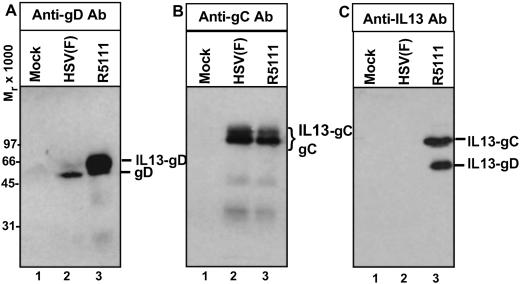
In the first stage of engineering a herpes simplex virus (HSV)-1 that specifically targets human malignant glioma cells, we constructed a recombinant virus designated R5111 in which we have ablated the binding sites for sulfated proteoglycans in glycoproteins B and C, replaced the amino-terminal 148 aa in glycoprotein C by IL-13 flanked at its amino terminus with a signal peptide, and inserted a second copy of IL-13 after the amino acid 24 of glycoprotein D. In the process, the binding site for HveA, a viral entry receptor, was disrupted. We have also transformed a cell line (J1.1) lacking HSV-1 receptors to express IL13Ralpha2 receptor (J13R cells). We report the following: the R5111 recombinant virus replicates as well as wild-type virus in a variety of cell lines including cell lines derived from brain tumors. R5111 failed to replicate in the parent J1.1 cell line but multiplied to titers similar to those obtained in other cell lines in the J13R cell line. On the basis of the evidence that R5111 can use the IL13Ralpha2 receptor for entry, we conclude that HSV-1 can use receptors other than HveA or nectins, provided it can bind to them. The domains of gD that interact with HveA and nectin receptors are independent of each other. Lastly, the fusogenic activities of the glycoproteins in the viral envelope are not dependent on a set of unique interactions between glycoprotein D and its receptor. The construction of R5111 opens the way for construction of viruses totally dependent on selected receptors for entry or imaging of targeted cells.
Figures


References
-
- Davis F. G., Freels, S., Grutsch, J., Barlas, S. & Brem, S. (1998) J. Neurosurg. 88, 1-10. - PubMed
-
- Pyles R. B., Warnick, R. E., Chalk, C. L., Szanti, B. E. & Parysek, L. M. (1997) Hum. Gene Ther. 8, 533-544. - PubMed
-
- Rampling R., Cruickshank, G., Papanastassiou, V., Nicoll, J., Hadley, D., Brennan, D., Petty, R., MacLean, A., Harland, J., McKie, E., et al. (2000) Gene Ther. 7, 859-866. - PubMed
-
- McKie E. A., Brown, S. M., MacLean, A. R. & Graham, D. I. (1998) Neuropathol. Appl. Neurobiol. 24, 367-372. - PubMed
-
- Markert J. M., Medlock, M. D., Rabkin, S. D., Gillespie, G. Y., Todo, T., Hunter, W. D., Palmer, C. A., Feigenbaum, F., Tornatore, C., Tufaro, F. & Martuza, R. L. (2000) Gene Ther. 7, 867-874. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
