

Microscopic evidence for a minus-end-directed power stroke in the kinesin motor ncd
- PMID: 12426369
- PMCID: PMC137211
- DOI: 10.1093/emboj/cdf622
Microscopic evidence for a minus-end-directed power stroke in the kinesin motor ncd
Abstract
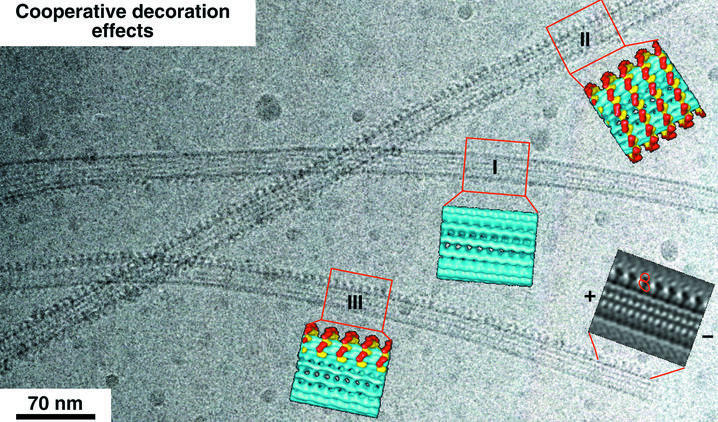
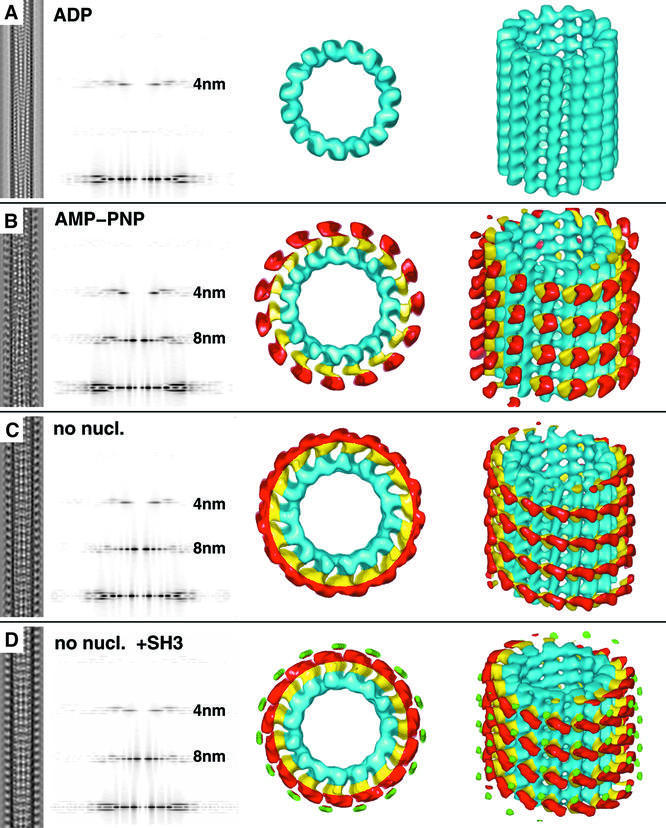
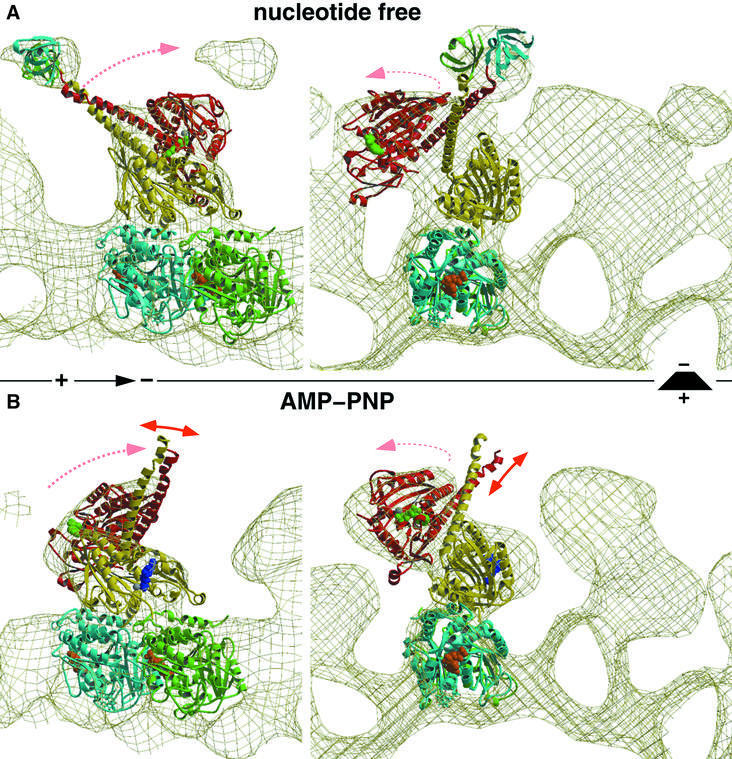
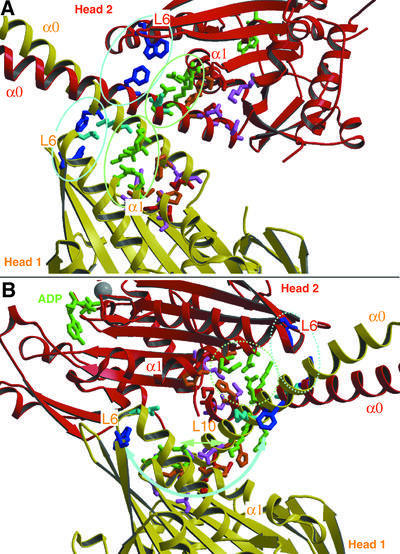
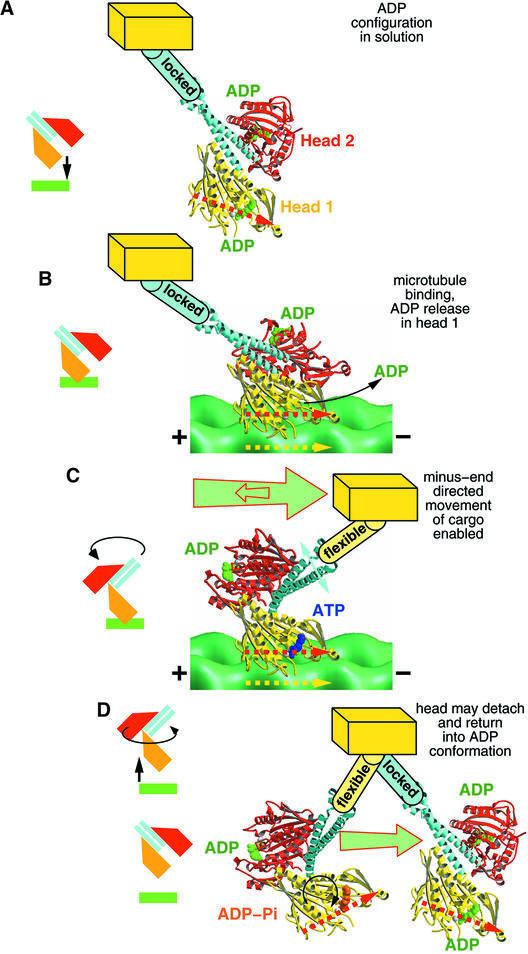
We used cryo-electron microscopy and image reconstruction to investigate the structure and microtubule-binding configurations of dimeric non-claret disjunctional (ncd) motor domains under various nucleotide conditions, and applied molecular docking using ncd's dimeric X-ray structure to generate a mechanistic model for force transduction. To visualize the alpha-helical coiled-coil neck better, we engineered an SH3 domain to the N-terminal end of our ncd construct (296-700). Ncd exhibits strikingly different nucleotide-dependent three-dimensional conformations and microtubule-binding patterns from those of conventional kinesin. In the absence of nucleotide, the neck adapts a configuration close to that found in the X-ray structure with stable interactions between the neck and motor core domain. Minus-end-directed movement is based mainly on two key events: (i) the stable neck-core interactions in ncd generate a binding geometry between motor and microtubule which places the motor ahead of its cargo in the minus-end direction; and (ii) after the uptake of ATP, the two heads rearrange their position relative to each other in a way that promotes a swing of the neck in the minus-end direction.
Figures
Similar articles
-
Structural comparison of dimeric Eg5, Neurospora kinesin (Nkin) and Ncd head-Nkin neck chimera with conventional kinesin.EMBO J. 2000 Oct 16;19(20):5308-14. doi: 10.1093/emboj/19.20.5308. EMBO J. 2000. PMID: 11032798 Free PMC article.
-
Motor domains of kinesin and ncd interact with microtubule protofilaments with the same binding geometry.J Mol Biol. 1997 Feb 7;265(5):553-64. doi: 10.1006/jmbi.1996.0757. J Mol Biol. 1997. PMID: 9048948
-
A new look at the microtubule binding patterns of dimeric kinesins.J Mol Biol. 2000 Apr 14;297(5):1087-103. doi: 10.1006/jmbi.2000.3627. J Mol Biol. 2000. PMID: 10764575
-
Kinesins and microtubules: their structures and motor mechanisms.Curr Opin Cell Biol. 2000 Feb;12(1):35-41. doi: 10.1016/s0955-0674(99)00054-x. Curr Opin Cell Biol. 2000. PMID: 10679355 Review.
-
Kinesin, 30 years later: Recent insights from structural studies.Protein Sci. 2015 Jul;24(7):1047-56. doi: 10.1002/pro.2697. Epub 2015 Jun 11. Protein Sci. 2015. PMID: 25975756 Free PMC article. Review.
Cited by
-
Decoration of microtubules in solution by the kinesin-14, Ncd.Acta Crystallogr D Biol Crystallogr. 2010 Nov;66(Pt 11):1218-23. doi: 10.1107/S0907444910021323. Epub 2010 Oct 20. Acta Crystallogr D Biol Crystallogr. 2010. PMID: 21041940 Free PMC article.
-
Regulation of KinI kinesin ATPase activity by binding to the microtubule lattice.J Cell Biol. 2003 Dec 8;163(5):963-71. doi: 10.1083/jcb.200304034. J Cell Biol. 2003. PMID: 14662742 Free PMC article.
-
Bound ion effects: Using machine learning method to study the kinesin Ncd's binding with microtubule.Biophys J. 2024 Sep 3;123(17):2740-2748. doi: 10.1016/j.bpj.2023.12.024. Epub 2023 Dec 30. Biophys J. 2024. PMID: 38160255 Free PMC article.
-
Bidirectional membrane tube dynamics driven by nonprocessive motors.Proc Natl Acad Sci U S A. 2008 Jun 10;105(23):7993-7. doi: 10.1073/pnas.0709677105. Epub 2008 Mar 10. Proc Natl Acad Sci U S A. 2008. PMID: 18332438 Free PMC article.
-
Neck rotation and neck mimic docking in the noncatalytic Kar3-associated protein Vik1.J Biol Chem. 2012 Nov 23;287(48):40292-301. doi: 10.1074/jbc.M112.416529. Epub 2012 Oct 7. J Biol Chem. 2012. PMID: 23043140 Free PMC article.
References
-
- Beuron F. and Hoenger,A. (2001) Structural analysis of the microtubule–kinesin complex by cryo-electron microscopy. In Vernos,I. (ed.), Methods in Molecular Biology. Humana Press, Totowa, NJ, pp. 235–254. - PubMed
-
- Case R.B., Pierce,D.W., Hom-Booher,N., Hart,C.L. and Vale,R.D. (1997) The directional preference of kinesin motors is specified by an element outside of the motor catalytic domain. Cell, 90, 959–966. - PubMed
-
- Chandra R., Salmon,E.D., Erickson,H.P., Lockhart,A. and Endow,S.A. (1993) Structural and functional domains of the Drosophila ncd microtubule motor protein. J. Biol. Chem., 268, 9005–9013. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
