

doi: 10.1093/emboj/cdf607.
RNA editing by ADARs is important for normal behavior in Caenorhabditis elegans
Affiliations
- PMID: 12426375
- PMCID: PMC137199
- DOI: 10.1093/emboj/cdf607
Item in Clipboard
RNA editing by ADARs is important for normal behavior in Caenorhabditis elegans
EMBO J.
.
Abstract
Here we take advantage of the well-characterized and simple nervous system of Caenorhabditis elegans to further our understanding of the functions of RNA editing. We describe the two C.elegans ADAR genes, adr-1 and adr-2, and characterize strains containing homozygous deletions in each, or both, of these genes. We find that adr-1 is expressed in most, if not all, cells of the C.elegans nervous system and also in the developing vulva. Using chemotaxis assays, we show that both ADARs are important for normal behavior. Biochemical, molecular and phenotypic analyses indicate that ADR-1 and ADR-2 have distinct roles in C.elegans, but sometimes act together.
Figures
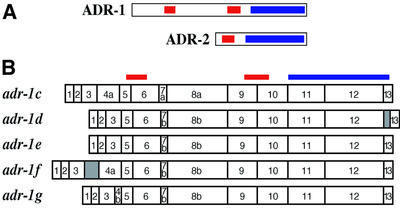
Fig. 1. Caenorhabditis elegans has two ADARs. (A) ADR-1 (adr-1c) and ADR-2 ORFs are shown, with boxes indicating dsRBMs (red) and the catalytic domain (blue). (B) Five splice forms of adr-1 are shown, with the relative positions of dsRBMs and catalytic domain indicated at the top. Notable features are as follows, with nucleotide positions given relative to the start of cosmid H15N15 (accession No. Z96100). The adr-1c cDNA (accession No. AY150815) includes exons 7a and 8a which are created by alternative 3′ splice sites (4877 and 4989 nucleotides). Compared with the b forms of these exons, 7a and 8a have nine and six additional nucleotides, respectively, and code for the addition of three amino acids (LLQ) or two amino acids (LQ), respectively. Exon 4 is absent from adr-1d and adr-1e cDNAs (accession Nos AY150816 and AY150817, respectively), resulting in a deletion of 66 amino acids that does not disrupt the ORF. adr-1d has an unspliced intron between exons 12 and 13 that inserts 19 amino acids into the catalytic domain near the C-terminus. adr-1f (accession No. AY150818) has an unspliced intron between exons 3 and 4, resulting in a premature stop in the intron, 301 nucleotides from the AUG codon. In adr-1g, an alternative 5′ splice site for intron 4 (4250 nucleotides) results in a 58 nucleotide exon 4b, truncated by 140 nucleotides compared with exon 4a; this predicts a frameshift and a premature stop codon in exon 5, 346 nucleotides from the 5′ end of adr-1g (accession No. AY150819). In theory, short forms of ADR-1 could be synthesized from adr-1f and adr-1g by using downstream methionines.
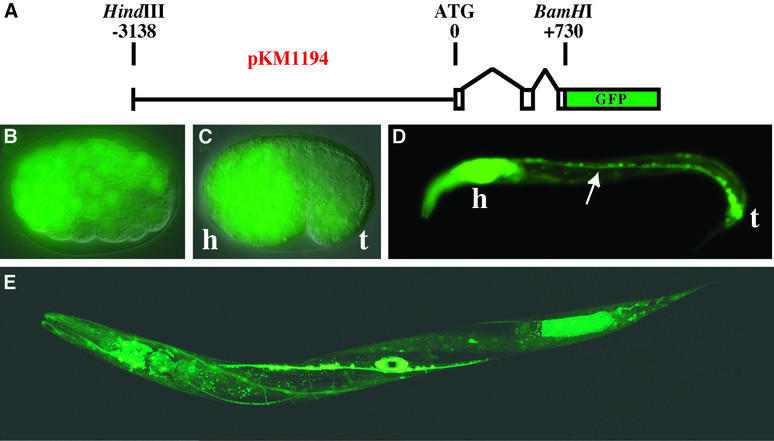
Fig. 2. Embryonic and larval expression of adr-1 in C.elegans. (A) The diagram shows the pKM1194 reporter construct used to generate adr-1::GFP expression lines (KM163–5). Nomarski/GFP merged images show that adr-1::GFP is expressed by late gastrulation (B, ≥100 cell stage), and by the comma stage (C) is predominantly neuronal (h, presumptive head; t, tail). Neuronal expression continues into larval stages as shown for an L1 worm (D), where expression was observed in the ventral nerve cord (arrow), along with intense expression in the head ganglia and nerve ring (h) and the tail neurons (t). A confocal image of a late L4 worm (E) shows expression in most cells of the nervous system, as well as in the developing vulva (see also Figure 3). Other tissues with weak expression include the pharynx and body wall muscle. We were unable to determine whether the expression in posterior intestinal cells is true adr-1 expression or an artifact of transgenic expression (Mello and Fire, 1995).
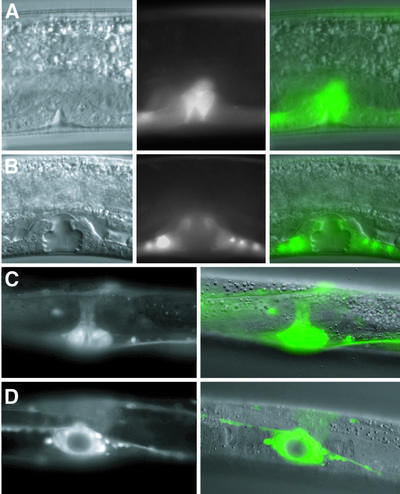
Fig. 3. adr-1 expression during vulva morphogenesis. Nomarski, GFP and merged images (left to right; KM165) show that adr-1::GFP is expressed in proliferating vulval precursor cells that are beginning to invaginate during late L3 (A), and continues to be localized to the developing vulva at the L4 Christmas tree stage (B). GFP and Nomarski/GFP merged images of a late L4 vulva (KM164) are shown with lateral (C) and ventral (D) views. All 22 cells derived from vulval precursor cells continue to express GFP into late L4. After the L4/adult molt, adr-1::GFP expression is no longer seen in the vulva.
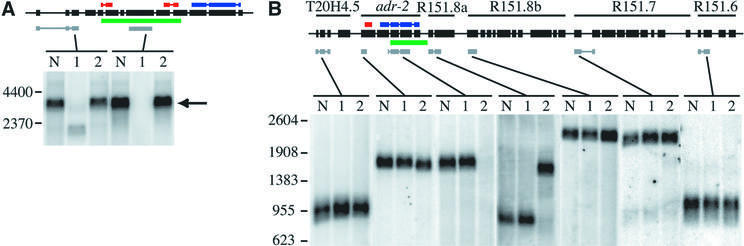
Fig. 4. Northern analyses of adr deletion strains. Black boxes with a line through the center represent the genomic structure of the adr-1 gene (A) or the six-gene adr-2 operon (B). Colored bars show the location of dsRBMs (red), catalytic domain (blue), sequences deleted in the mutants (green) and the regions of cDNA sequences used to probe northerns (gray). Gels were loaded with poly(A)+ RNA from three C.elegans strains: wild-type (lane N), adr-1(gv6) (lane 1) and adr-2(gv42) (lane 2). (A) Hybridization with a probe to the 5′ region of adr-1 shows a band corresponding to the full-length transcript (arrow) in wild-type and adr-2(gv42) samples and a shorter transcript in the adr-1(gv6) deletion strain; the strong hybridization signal of this experiment did not allow resolution of alternative splice forms. A probe to sequences within the deleted region shows equivalent expression in wild-type and adr-2(gv42) lanes and no signal in the adr-1(gv6) lane. (B) Hybridization with probes to various regions of the six-gene operon shows that transcripts from T20H4.5, R151.8b, R151.7 and R151.6 are unaffected by the deletion in adr-2(gv42); all genes are named as previously specified (Hough et al., 1999). A probe to the 5′ region of adr-2 hybridizes to a slightly shorter transcript in the adr-2(gv42) deletion strain, while a probe within the deleted region shows normal hybridization to wild-type and adr-1(gv6) RNA but no signal in the adr-2(gv42) RNA. A probe to R151.8a, the gene just downstream of adr-2, confirms that adr-2(gv42) contains a chimeric transcript that fuses adr-2 to R151.8a. The chimeric RNA is predicted to result in a chimeric protein with the first two exons of adr-2 in-frame with the protein encoded by R151.8a. An RT–PCR product spanning the junction of the adr-2::R151.8a fusion was cloned and sequenced, and confirmed results of the northern analyses.
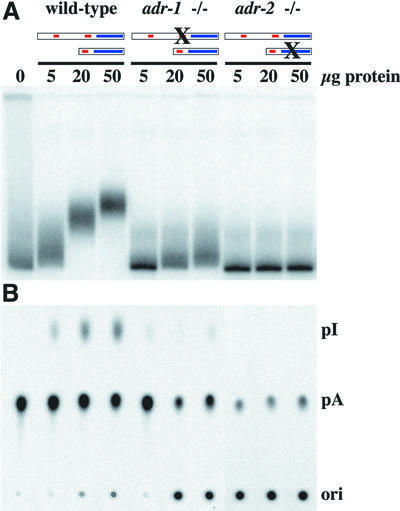
Fig. 5. In vitro assays of deaminase activity in wild-type and mutant worms. 32P-labeled dsRNA was incubated with various amounts of protein extract, for 2 h at 20°C. In (A), reaction products were electrophoresed on a native gel, and a phosphorimage of the gel is shown. This assay is based on the fact that adenosine deamination changes the structure of a dsRNA and thus its mobility on a native gel. In (B), the conversion of [32P]AMP to [32P]IMP was measured directly by TLC. After incubation, RNA was isolated, digested to mononucleotides and chromatographed on a TLC plate; a phosphor image of the TLC plate is shown. Ori, origin; pI, [32P]IMP; pA, [32P]AMP.
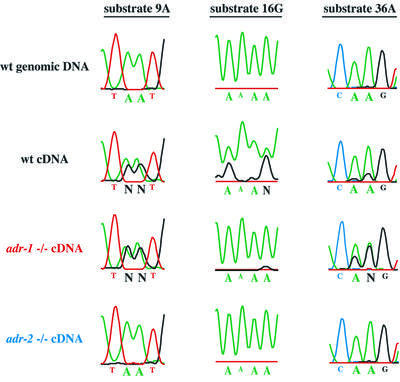
Fig. 6. Analysis of editing in RNA isolated from wild-type and mutant worms. The figure shows regions of electropherograms generated by sequencing PCR products amplified from genomic DNA, wild-type cDNA or adr mutant cDNAs, for three different ADAR substrates. In genomic DNA (top line), the sites indicated by large type are exclusively adenosines (green peaks) but, in wild-type cDNAs (second line), they are a mixture of adenosines (green peaks) and guanosines (black peaks), indicating that these sites are edited. The relative area under the peaks is a measure of the efficiency of editing. Editing sites shown for each substrate were chosen to illustrate three different effects of the adr-1 deletion and are not necessarily representative of all editing sites in these substrates. N indicates sites with nearly equal amounts of A and G that could not be called by the sequencing software. Note that the failure of the A trace of substrate 16G to go to baseline is typical of regions of homopolymer sequence; in other experiments, editing at these sites was confirmed by sequencing individual cDNA clones (Morse and Bass, 1999).
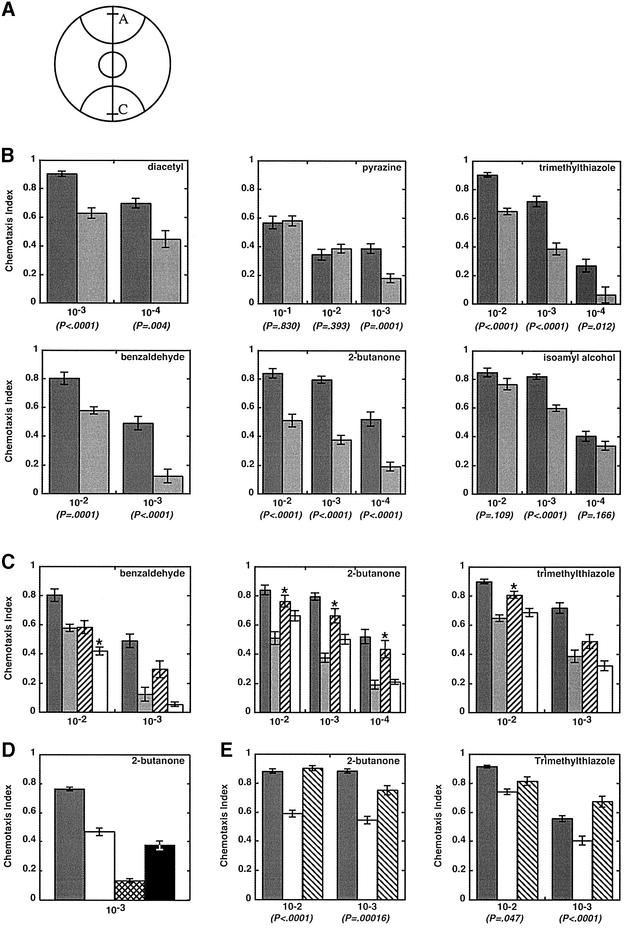
Fig. 7. Chemotaxis population assays of wild-type and mutant animals. (A) Plates were marked with semi-circles (radius, 28 mm) to delineate attractant (A) and control (C) regions and spotted with 1 µl of attractant 5 mm from one edge (line in region A) and 1 µl of ethanol 5 mm from the opposite edge (line in region C); 1 µl of sodium azide (1 M) was placed at the same position to anesthetize animals reaching those areas. Between 100 and 200 adult animals (day 2) were placed in a circle at the center of the plate. After 2 h, animals were counted to calculate a chemotaxis index: [animals at (A) – animals at (C)]/[total animals on plate]. (B) Bar graphs show data from assays as in (A) for wild-type animals (dark gray) and adr-1;adr-2 double mutants (light gray). Chemotaxis index values are the average of ≥10 independent determinations, and error bars indicate the SEM; P-values show the t-probability from an unpaired (non-directional) two-tailed test. Attractants were diluted in ethanol as specified on the x-axis (v/v), except for pyrazine, which is a solid and was dissolved in ethanol (w/v). Chemotaxis values observed with wild-type animals all matched reported values, except for pyrazine (Bargmann et al., 1993), possibly because pyrazine had to be dissolved, increasing the chance of differences between laboratories. (C) As in (B) except that the wild type (dark gray) and adr-1;adr-2 double mutants (light gray) were compared with the single mutants, adr-1(gv6) (hatched) and adr-2(gv42) (open). Asterisks mark data for the single mutants that were significantly different from the double mutant at P < 0.01. (D) Chemotaxis to 2-butanone (10–3) was compared for wild-type (dark gray), adr-2(gv42) (open), che-2(e1033) (cross hatched) and odr-3(n2150) (black) animals. Chemotaxis values are averages of ≥7 independent determinations; error bars show the SEM. che-2(e1033) (P = 0.0001), but not odr-3(n2150) (P = 0.03), chemotaxis data were significantly different from those of adr-2(gv42). (E) Chemotaxis assays comparing the wild type (dark gray) and adr-2(gv42) (open) with the adr-2(gv42) rescue line B (hatched). P-values show that adr-2(gv42) rescue worms are significantly different from adr-2(gv42), except at the 10–2 dilution of trimethylthiazole. The adr-2(gv42) rescue lines were assayed by counting only the GFP-expressing portion of the population.
References
-
- Anderson P. (1995) Mutagenesis. In Epstein,H. and Shakes,D. (eds), Caenorhabditis elegans: Modern Biological Analysis of an Organism. Vol. 48. Academic Press, San Diego, CA, pp. 31–58.
-
- Baltimore D. (2001) Our genome unveiled. Nature, 409, 814–816. - PubMed
-
- Bargmann C.I. and Kaplan,J.M. (1998) Signal transduction in the Caenorhabditis elegans nervous system. Annu. Rev. Neurosci., 21, 279–308. - PubMed
-
- Bargmann C.I., Hartwieg,E. and Horvitz,H.R. (1993) Odorant-selective genes and neurons mediate olfaction in C.elegans. Cell, 74, 515–527. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
