

Mechanisms of amygdala modulation of hippocampal plasticity
- PMID: 12427848
- PMCID: PMC6757810
- DOI: 10.1523/JNEUROSCI.22-22-09912.2002
Mechanisms of amygdala modulation of hippocampal plasticity
Abstract
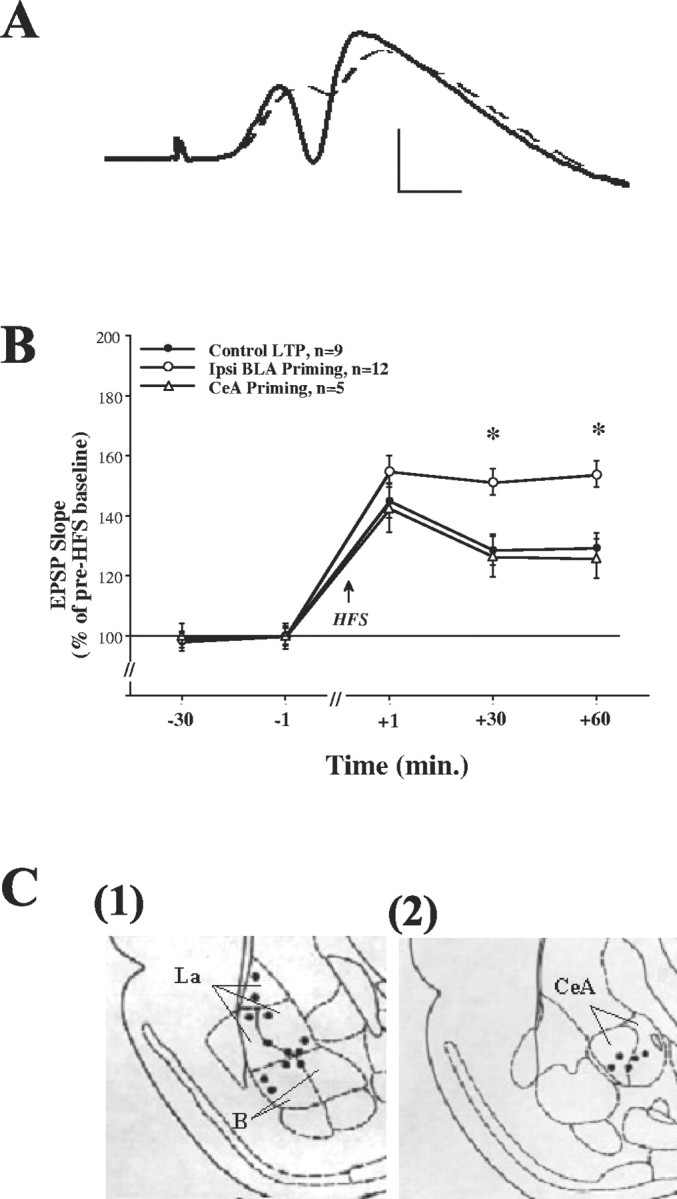
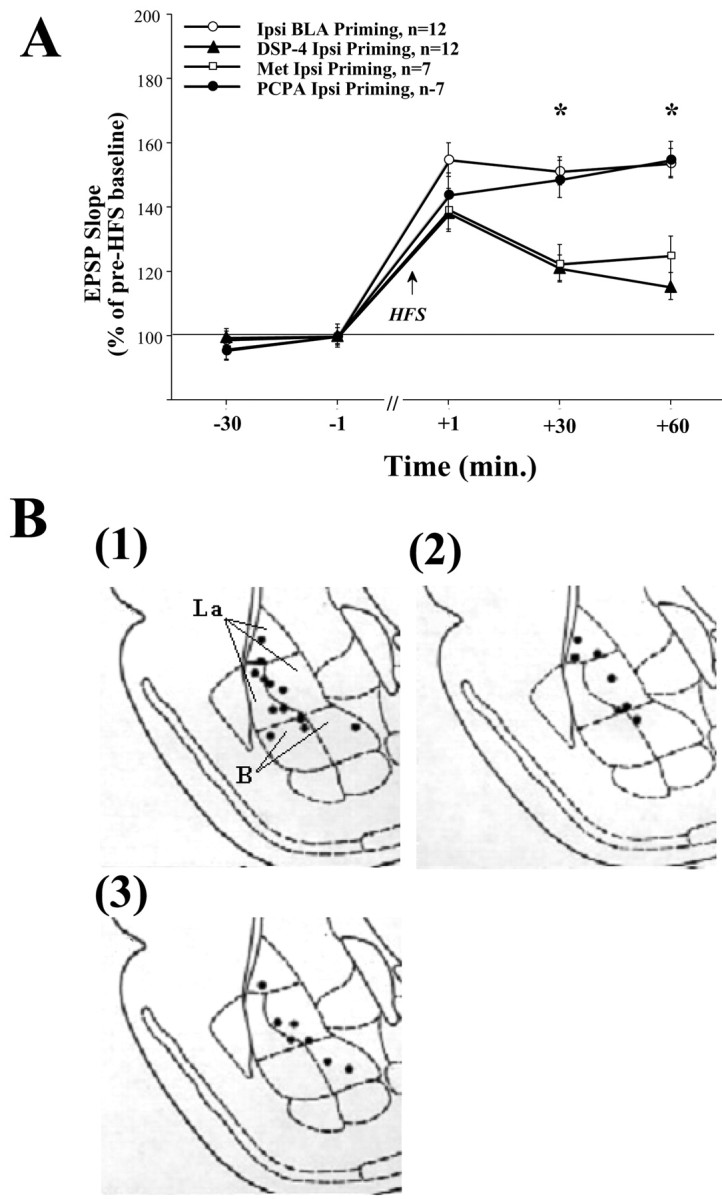
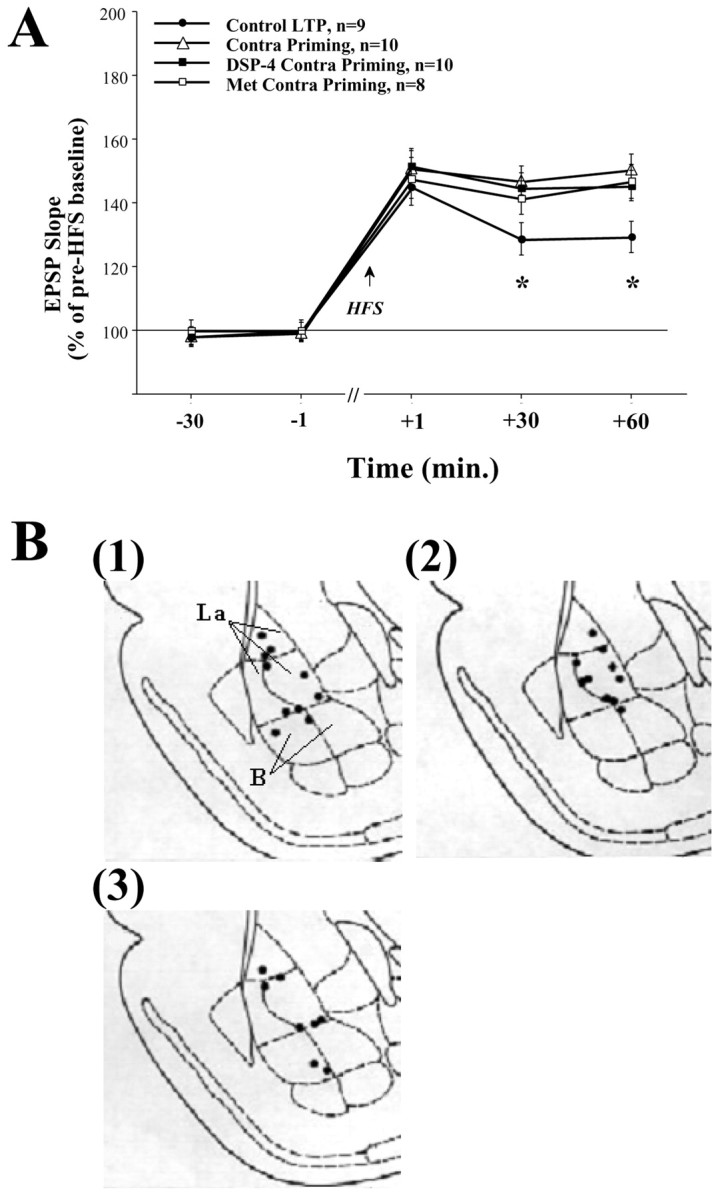
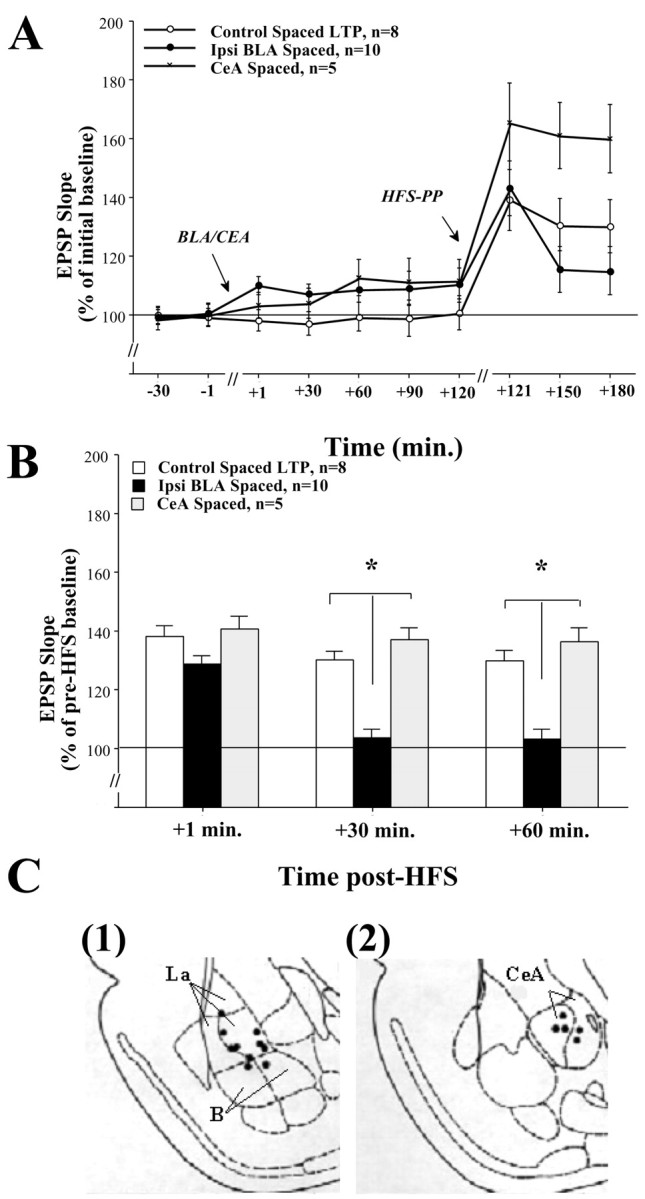
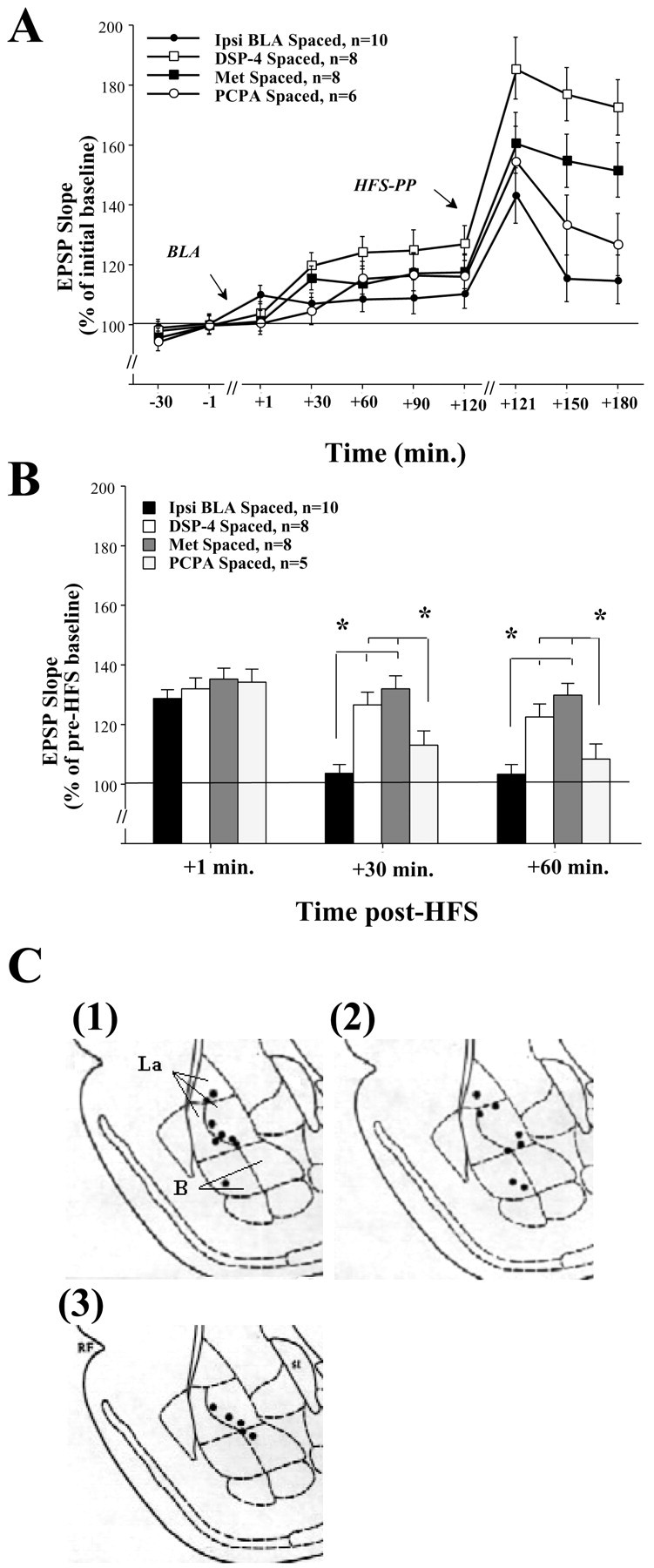
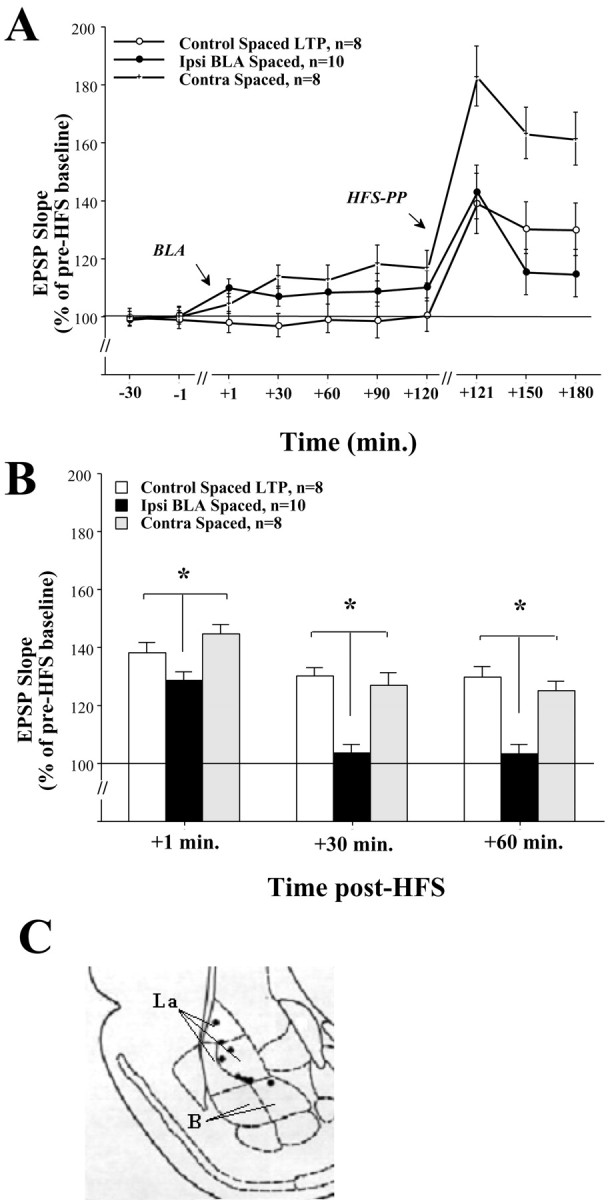
Basolateral amygdala (BLA) activation by emotional arousal modulates memory-related processes in the hippocampus. We have shown (Akirav and Richter-Levin, 1999b) that activating the BLA before perforant path (PP) tetanization has a biphasic effect on hippocampal plasticity; priming the BLA immediately before PP tetanization results in the enhancement of dentate gyrus (DG) long-term potentiation (LTP) (an "emotional tag"), whereas stimulation in a spaced interval results in the suppression of DG-LTP. Here, we aimed to elucidate the mechanisms underlying BLA modulation of DG-LTP and specifically to examine whether the stress hormones norepinephrine (NE) and corticosterone (CORT) are main mediators of the BLA biphasic effects. We found that the BLA affects hippocampal plasticity in a complex manner; BLA priming enhanced DG-LTP, and both NE and CORT mediated this effect. Furthermore, we found that ipsilateral BLA spaced activation (2 hr before PP tetanization) suppressed DG-LTP and that this suppressive effect was also mediated by NE and CORT. Priming the contralateral BLA enhanced DG-LTP similarly to the ipsilateral enhancement, but neither NE nor CORT mediated this effect. The spaced activation of the contralateral BLA did not suppress DG-LTP. Taken together, these results suggest that differential mechanisms underlie the ipsilateral and contralateral BLA effects on hippocampal plasticity. Hence, the BLA modulates hippocampal memory processes, presumably via the mediation of the stress hormones NE and CORT, to establish a diverse memory of the experience. Possibly, at the onset of an emotional event the stress hormones permissively mediate plasticity. However, their prolonged presence in the system may suppress the cognitive response to stress.
Figures
References
-
- Abe K. Modulation of hippocampal long-term potentiation by the amygdala: a synaptic mechanism linking emotion and memory. Jpn J Pharmacol. 2001;86:18–22. - PubMed
-
- Akirav I, Richter-Levin G. Priming stimulation in the basolateral amygdala modulates synaptic plasticity in the rat dentate gyrus. Neurosci Lett. 1999a;30:83–86. - PubMed
-
- Amorapanth P, LeDoux JE, Nader K. Different lateral amygdala outputs mediate reactions and actions elicited by a fear-arousing stimulus. Nat Neurosci. 2000;3:74–79. - PubMed
-
- Bhatnagar S, Costall B, Smythe JW. Hippocampal cholinergic blockade enhances hypothalamic-pituitary-adrenal responses to stress. Brain Res. 1997;22:244–248. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous