

Characterization of AtCDC48. Evidence for multiple membrane fusion mechanisms at the plane of cell division in plants
- PMID: 12427991
- PMCID: PMC166645
- DOI: 10.1104/pp.011742
Characterization of AtCDC48. Evidence for multiple membrane fusion mechanisms at the plane of cell division in plants
Abstract
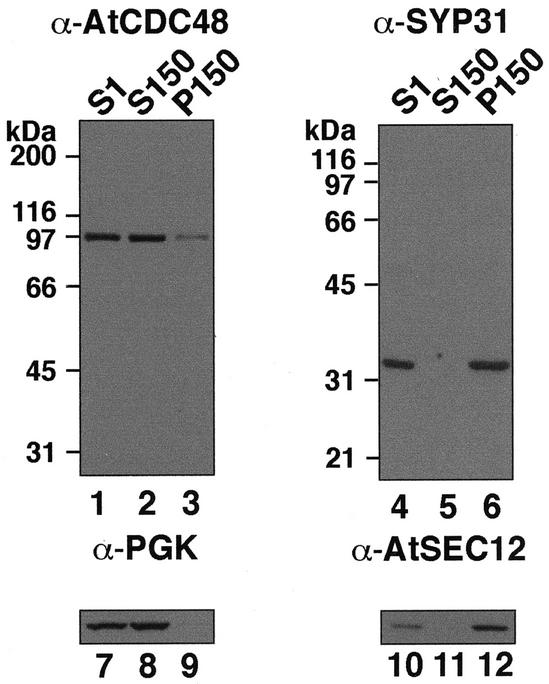
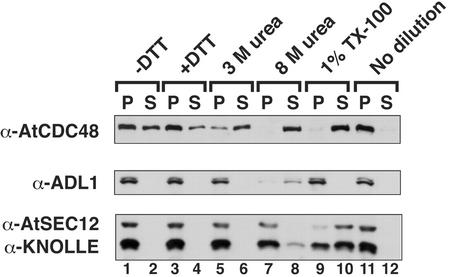
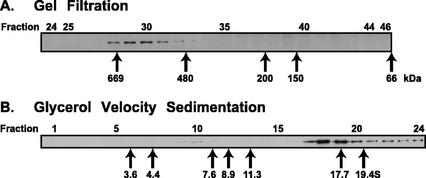
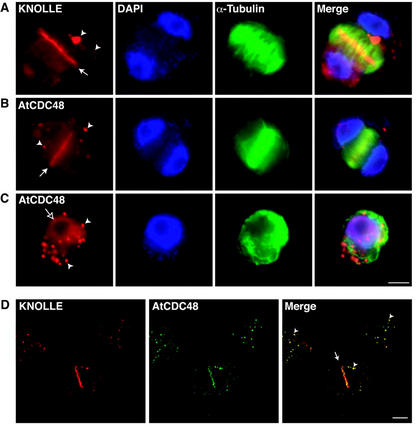
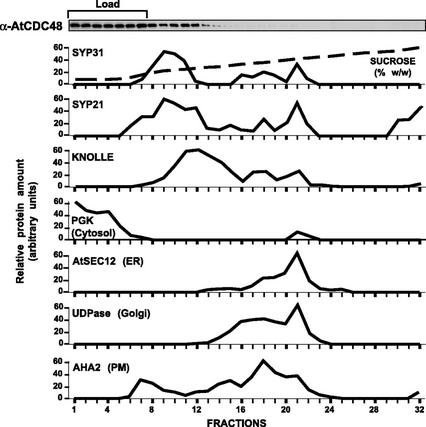
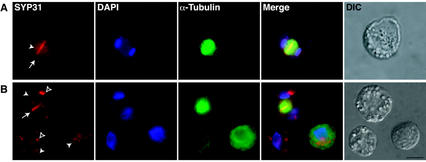
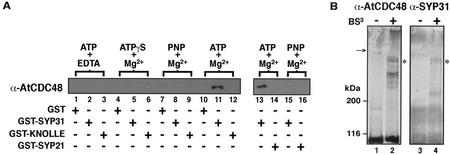
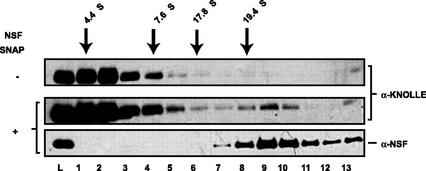
The components of the cellular machinery that accomplish the various complex and dynamic membrane fusion events that occur at the division plane during plant cytokinesis, including assembly of the cell plate, are not fully understood. The most well-characterized component, KNOLLE, a cell plate-specific soluble N-ethylmaleimide-sensitive fusion protein (NSF)-attachment protein receptor (SNARE), is a membrane fusion machine component required for plant cytokinesis. Here, we show the plant ortholog of Cdc48p/p97, AtCDC48, colocalizes at the division plane in dividing Arabidopsis cells with KNOLLE and another SNARE, the plant ortholog of syntaxin 5, SYP31. In contrast to KNOLLE, SYP31 resides in defined punctate membrane structures during interphase and is targeted during cytokinesis to the division plane. In vitro-binding studies demonstrate that AtCDC48 specifically interacts in an ATP-dependent manner with SYP31 but not with KNOLLE. In contrast, we show that KNOLLE assembles in vitro into a large approximately 20S complex in an Sec18p/NSF-dependent manner. These results suggest that there are at least two distinct membrane fusion pathways involving Cdc48p/p97 and Sec18p/NSF that operate at the division plane to mediate plant cytokinesis. Models for the role of AtCDC48 and SYP31 at the division plane will be discussed.
Figures
References
-
- Acharya U, Jacobs R, Peters JM, Watson N, Farquhar MG, Malhotra V. The formation of Golgi stacks from vesiculated Golgi membranes requires two distinct fusion events. Cell. 1995;82:895–904. - PubMed
-
- Aguilar RM, Bustamante JJ, Hernandez PG, Martinez AO, Haro LS. Precipitation of dilute chromatographic samples (ng/ml) containing interfering substances for SDS-PAGE. Anal Biochem. 1999;267:344–350. - PubMed
-
- Assaad FF, Mayer U, Wanner G, Jürgens G. The KEULE gene is involved in cytokinesis in Arabidopsis. Mol Gen Genet. 1996;253:267–277. - PubMed
-
- Banfield DK, Lewis MJ, Pelham HR. A SNARE-like protein required for traffic through the Golgi complex. Nature. 1995;375:806–809. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
