

Control of intracellular chloride concentration and GABA response polarity in rat retinal ON bipolar cells
- PMID: 12433959
- PMCID: PMC2290660
- DOI: 10.1113/jphysiol.2002.024877
Control of intracellular chloride concentration and GABA response polarity in rat retinal ON bipolar cells
Abstract
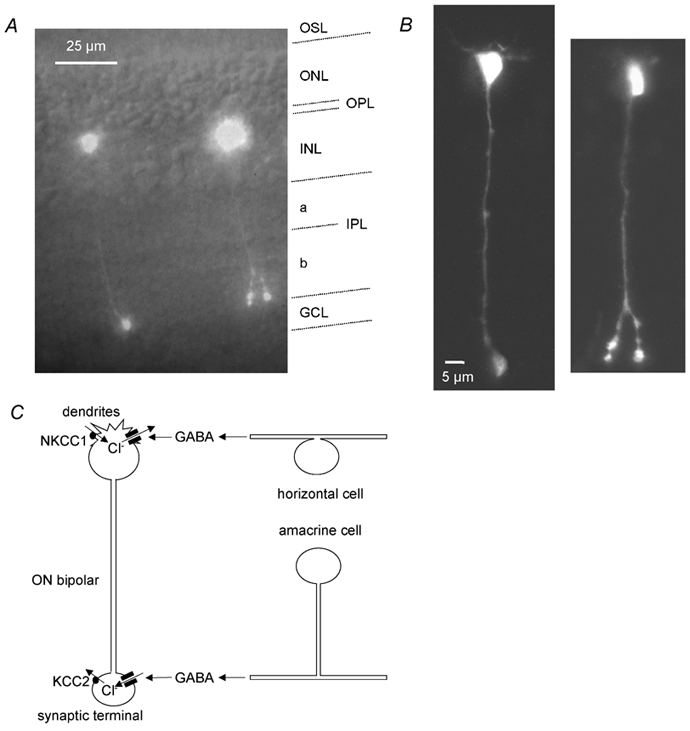
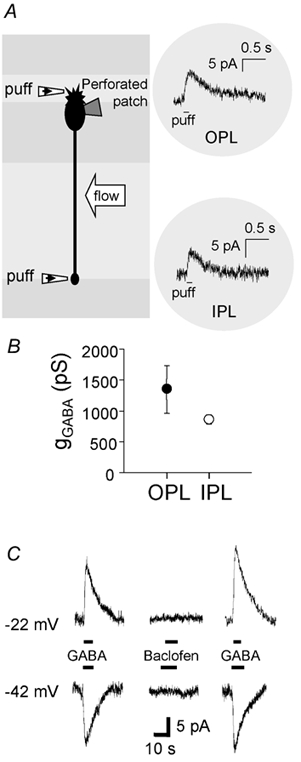
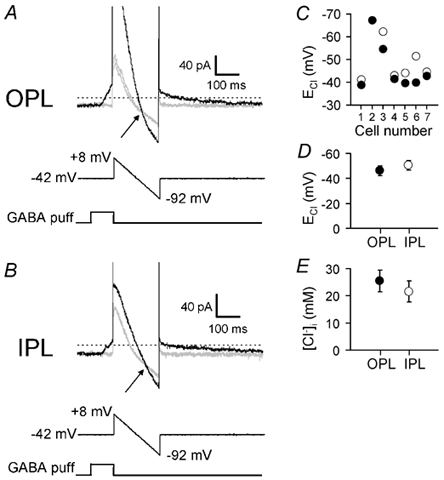
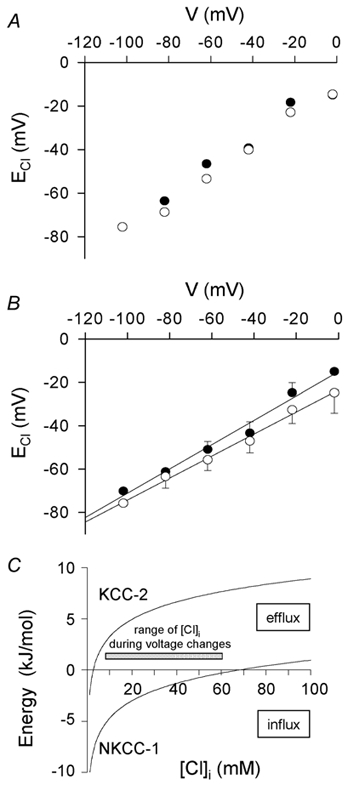
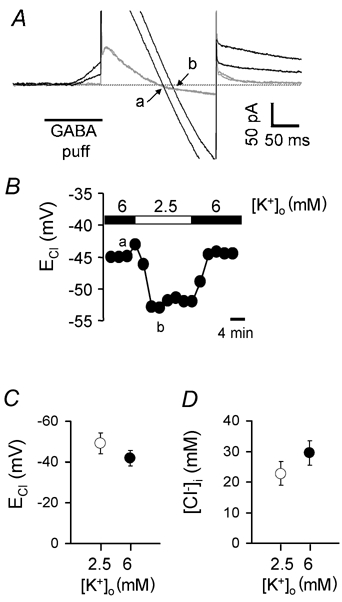
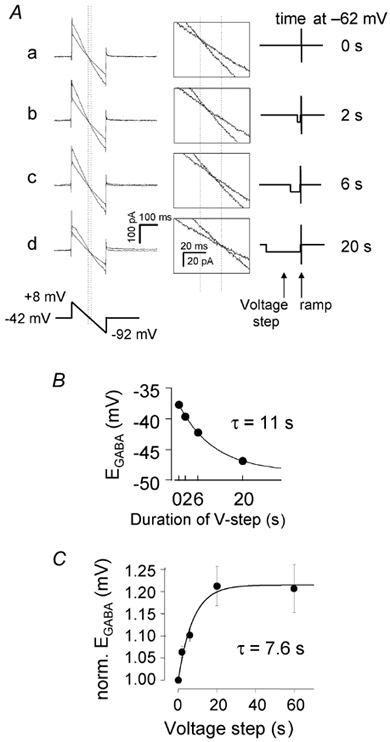
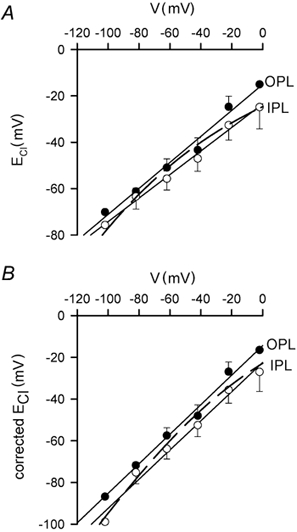
GABAergic modulation of retinal bipolar cells plays a crucial role in early visual processing. It helps to form centre-surround receptive fields which filter the visual signal spatially at the bipolar cell dendrites in the outer retina, and it produces temporal filtering at the bipolar cell synaptic terminals in the inner retina. The observed chloride transporter distribution in ON bipolar cells has been predicted to produce an intracellular chloride concentration, [Cl(-)](i), that is significantly higher in the dendrites than in the synaptic terminals. This would allow dendritic GABA-gated Cl(-) channels to generate the depolarization needed for forming the lateral inhibitory surround of the cell's receptive field, while synaptic terminal GABA-gated Cl(-) channels generate the hyperpolarization needed for temporal shaping of the light response. In contrast to this idea, we show here that in ON bipolar cells [Cl(-)](i) is only slightly higher in the dendrites than in the synaptic terminals, and that GABA-gated channels in the dendrites may generate a hyperpolarization rather than a depolarization. We also show that [Cl(-)](i) is controlled by movement of Cl(-) through ion channels in addition to transporters, that changes of [K(+)](o) alter [Cl(-)](i) and that voltage-dependent equilibration of [Cl(-)](i) in bipolar cells will produce a time-dependent adaptation of GABAergic modulation with a time constant of 8 s after illumination-evoked changes of membrane potential. Time-dependent adaptation of [Cl(-)](i) to voltage changes in retinal bipolar cells may add a previously unsuspected layer of temporal processing to signals as they pass through the retina.
Figures
References
-
- Akaike N. Gramicidin perforated patch recording and intracellular chloride activity in excitable cells. Progress in Biophysics and Molecular Biology. 1996;65:251–264. - PubMed
-
- Ben-Ari Y, Tseeb V, Raggozzino D, Khazipov R, Gaiarsa JL. γ-Aminobutyric acid (GABA), a fast excitatory transmitter which may regulate the development of hippocampal neurones in early postnatal life. Progress in Brain Research. 1994;102:261–273. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous