

Mitogen-activated protein kinase signaling in postgermination arrest of development by abscisic acid
- PMID: 12434021
- PMCID: PMC137798
- DOI: 10.1073/pnas.242607499
Mitogen-activated protein kinase signaling in postgermination arrest of development by abscisic acid
Abstract
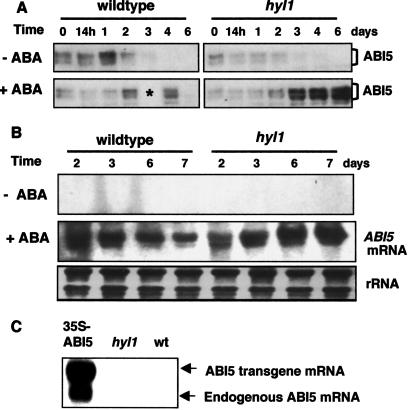
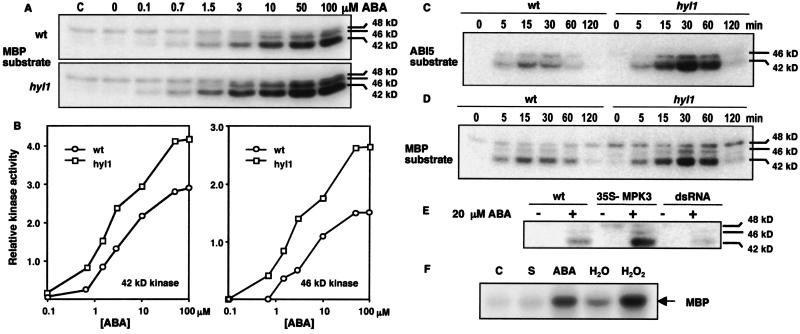
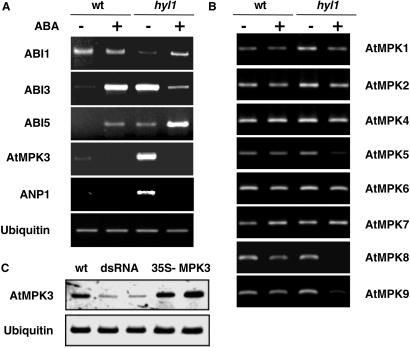
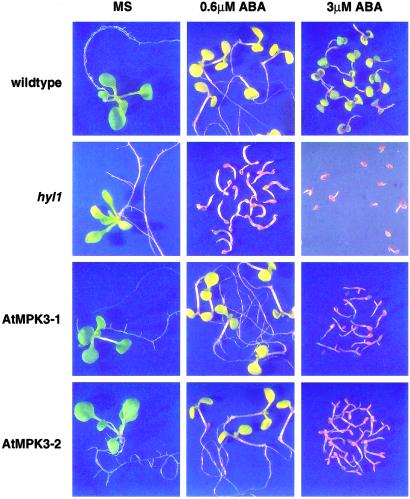
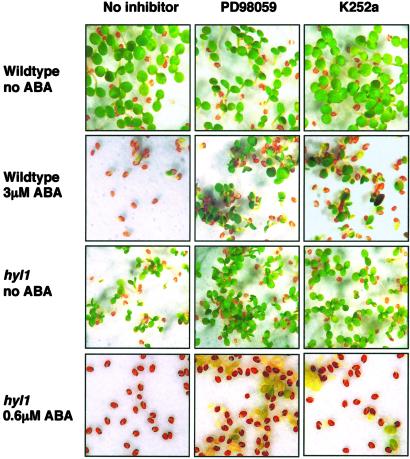
Abscisic acid (ABA) mediates plant responses to environmental stress, particularly to water status. During germination, the embryo emerges from dormancy as the ABA concentration declines. Exposure to exogenous ABA during germination arrests development rapidly, but reversibly, enabling seedlings to withstand early water stress without loss of viability. Postgermination proteolytic degradation of the essential ABI5 transcription factor is interrupted by perception of an increase in ABA concentration, leading to ABI5 accumulation and reactivation of embryonic genes. Making use of the ABA-hypersensitive hyl1 mutant of Arabidopsis, we show that the ABA signal is transmitted to the transcriptional apparatus through mitogen-activated protein kinase signaling.
Figures
Similar articles
-
A postgermination developmental arrest checkpoint is mediated by abscisic acid and requires the ABI5 transcription factor in Arabidopsis.Proc Natl Acad Sci U S A. 2001 Apr 10;98(8):4782-7. doi: 10.1073/pnas.081594298. Epub 2001 Apr 3. Proc Natl Acad Sci U S A. 2001. PMID: 11287670 Free PMC article.
-
Arabidopsis RAV1 transcription factor, phosphorylated by SnRK2 kinases, regulates the expression of ABI3, ABI4, and ABI5 during seed germination and early seedling development.Plant J. 2014 Nov;80(4):654-68. doi: 10.1111/tpj.12670. Plant J. 2014. PMID: 25231920
-
Isolation and characterization of novel mutants affecting the abscisic acid sensitivity of Arabidopsis germination and seedling growth.Plant Cell Physiol. 2004 Oct;45(10):1485-99. doi: 10.1093/pcp/pch171. Plant Cell Physiol. 2004. PMID: 15564532
-
Light and abscisic acid interplay in early seedling development.New Phytol. 2021 Jan;229(2):763-769. doi: 10.1111/nph.16963. Epub 2020 Oct 25. New Phytol. 2021. PMID: 32984965 Review.
-
Impacts of altered RNA metabolism on abscisic acid signaling.Curr Opin Plant Biol. 2003 Oct;6(5):463-9. doi: 10.1016/s1369-5266(03)00084-0. Curr Opin Plant Biol. 2003. PMID: 12972047 Review.
Cited by
-
Genome-Wide Identification and Expression Analysis of the Mitogen-Activated Protein Kinase Gene Family in Cassava.Front Plant Sci. 2016 Aug 30;7:1294. doi: 10.3389/fpls.2016.01294. eCollection 2016. Front Plant Sci. 2016. PMID: 27625666 Free PMC article.
-
Quantitative phosphoproteomics identifies SnRK2 protein kinase substrates and reveals the effectors of abscisic acid action.Proc Natl Acad Sci U S A. 2013 Jul 2;110(27):11205-10. doi: 10.1073/pnas.1308974110. Epub 2013 Jun 17. Proc Natl Acad Sci U S A. 2013. PMID: 23776212 Free PMC article.
-
Evidence of a role for tyrosine dephosphorylation in the control of postgermination arrest of development by abscisic acid in Arabidopsis thaliana L.Planta. 2006 Jan;223(2):381-5. doi: 10.1007/s00425-005-0135-6. Epub 2005 Oct 7. Planta. 2006. PMID: 16211388
-
Arabidopsis TARGET OF RAPAMYCIN interacts with RAPTOR, which regulates the activity of S6 kinase in response to osmotic stress signals.Plant Cell. 2006 Feb;18(2):477-90. doi: 10.1105/tpc.105.035931. Epub 2005 Dec 23. Plant Cell. 2006. PMID: 16377759 Free PMC article.
-
A Method for Assaying of Protein Kinase Activity In Vivo and Its Use in Studies of Signal Transduction in Strawberry Fruit Ripening.Int J Mol Sci. 2021 Sep 28;22(19):10495. doi: 10.3390/ijms221910495. Int J Mol Sci. 2021. PMID: 34638834 Free PMC article.
References
-
- Assmann S. M. & Wang, X.-Q. (2001) Curr. Opin. Plant Biol. 4 421-428. - PubMed
-
- Hetherington A. M. (2001) Cell 107 711-714. - PubMed
-
- Bewley J. D. & Black, M., (1994) Seeds: Physiology of Development and Germination (Plenum, New York).
-
- Rock C. D. & Quatrano, R. S. (1995) in Plant Hormones: Physiology, Biochemistry and Molecular Biology, ed. Davies, P. J. (Kluwer Academic, London), pp. 671–697.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
