

GapA and CrmA coexpression is essential for Mycoplasma gallisepticum cytadherence and virulence
- PMID: 12438360
- PMCID: PMC133084
- DOI: 10.1128/IAI.70.12.6839-6845.2002
GapA and CrmA coexpression is essential for Mycoplasma gallisepticum cytadherence and virulence
Abstract
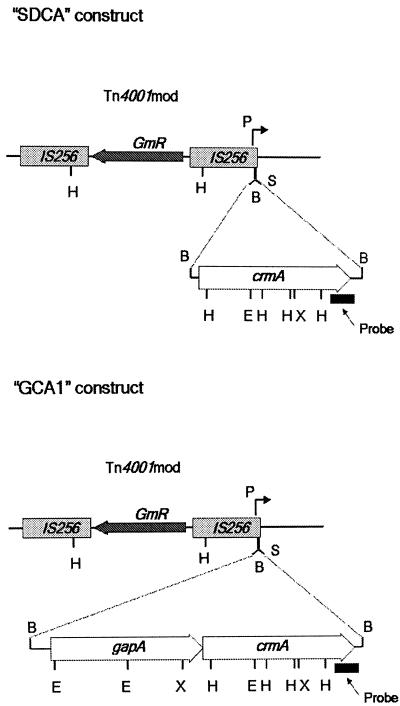
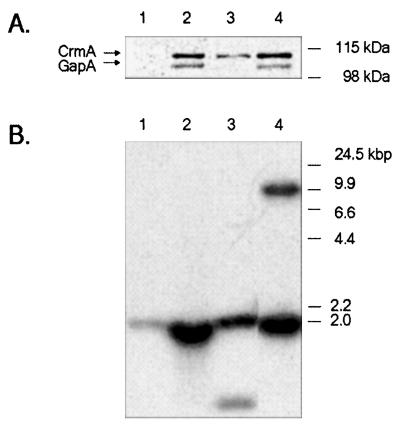
It was previously demonstrated that avirulent Mycoplasma gallisepticum strain R(high) (passage 164) is lacking three proteins that are expressed in its virulent progenitor, strain R(low) (passage 15). These proteins were identified as the cytadhesin molecule GapA, the putative cytadhesin-related molecule CrmA, and a component of a high-affinity transporter system, HatA. Complementation of R(high) with wild-type gapA restored expression in the transformant (GT5) but did not restore the cytadherence phenotype and maintained avirulence in chickens. These results suggested that CrmA might play an essential role in the M. gallisepticum cytadherence process. CrmA is encoded by the second gene in the gapA operon and shares significant sequence homology to the ORF6 gene of Mycoplasma pneumoniae, which has been shown to play an accessory role in the cytadherence process. Complementation of R(high) with wild-type crmA resulted in the transformant (SDCA) that lacked the cytadherence and virulence phenotype comparable to that found in R(high) and GT5. In contrast, complementation of R(high) with the entire wild-type gapA operon resulted in the transformant (GCA1) that restored cytadherence to the level found in wild-type R(low). In vivo pathogenesis trials revealed that GCA1 had regained virulence, causing airsacculitis in chickens. These results demonstrate that both GapA and CrmA are required for M. gallisepticum cytadherence and pathogenesis.
Figures
 ], transmembrane region (TM); [
], transmembrane region (TM); [ ], putative signal peptidase cleavage site; [
], putative signal peptidase cleavage site; [ ], domains shared by carbohydrate binding proteins (determined by performing a BLOCKS database search [26]); [▿], heparin sulfate binding-like domains (determined by performing a FastA search [58] with data from references , , , , and 70); [▴], lectin-like recognition domains (determined by performing a Patternmatch search with data from references and 67); [◊], other domains shared by some adhesion molecules (determined by performing a RPS-BLAST search on the Conserved Domain database [2]); [□], interaction domains shared proteins involved in the signal transduction-MARCKS family (determined by performing a BLOCKS database search [26]); [], histone H1, HMG14/HMG17 signature (GapA), H2B and H5 signature, HMG14/HMG17 family, DNA polymerase family X, and AlgR homology (CrmA) (determined by performing a BLOCKS database search [26]). [∗], ATP/GTP-binding site motif A (P-loop) (determined by performing a PROSITE motif search [11]).
], domains shared by carbohydrate binding proteins (determined by performing a BLOCKS database search [26]); [▿], heparin sulfate binding-like domains (determined by performing a FastA search [58] with data from references , , , , and 70); [▴], lectin-like recognition domains (determined by performing a Patternmatch search with data from references and 67); [◊], other domains shared by some adhesion molecules (determined by performing a RPS-BLAST search on the Conserved Domain database [2]); [□], interaction domains shared proteins involved in the signal transduction-MARCKS family (determined by performing a BLOCKS database search [26]); [], histone H1, HMG14/HMG17 signature (GapA), H2B and H5 signature, HMG14/HMG17 family, DNA polymerase family X, and AlgR homology (CrmA) (determined by performing a BLOCKS database search [26]). [∗], ATP/GTP-binding site motif A (P-loop) (determined by performing a PROSITE motif search [11]).References
-
- Alexandrov, N. N., R. Nussinov, and R. M. Zimmer. 1996. Fast protein fold recognition via sequence to structure alignment and contact capacity potentials, p. 53-72. In L. Hunter and T. E. Klein (ed.), Pacific symposium on biocomputing ’96, World Scientific Publishing Co., Singapore. - PubMed
-
- Baldi, P., S. Brunak, P. Frasconi, G. Soda, and G. Pollastri. 1999. Exploiting the past and the future in protein secondary structure prediction. Bioinformatics 15:937-946. - PubMed
-
- Bates, P. A., L. A. Kelley, R. M. MacCallum, and M. J. Sternberg. 2001. Enhancement of protein modeling by human intervention in applying the automatic programs 3D-JIGSAW and 3D-PSSM. Proteins 45:39-46. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
