

doi: 10.1083/jcb.200204065.
Epub 2002 Nov 18.
Activation of myosin V-based motility and F-actin-dependent network formation of endoplasmic reticulum during mitosis
Affiliations
- PMID: 12438410
- PMCID: PMC2173107
- DOI: 10.1083/jcb.200204065
Item in Clipboard
Activation of myosin V-based motility and F-actin-dependent network formation of endoplasmic reticulum during mitosis
J Cell Biol.
.
Abstract
It is widely believed that microtubule- and F-actin-based transport of cytoplasmic organelles and membrane fusion is down-regulated during mitosis. Here we show that during the transition of Xenopus egg extracts from interphase to metaphase myosin V-driven movement of small globular vesicles along F-actin is strongly inhibited. In contrast, the movement of ER and ER network formation on F-actin is up-regulated in metaphase extracts. Our data demonstrate that myosin V-driven motility of distinct organelles is differently controlled during the cell cycle and suggest an active role of F-actin in partitioning, positioning, and membrane fusion of the ER during cell division.
Figures
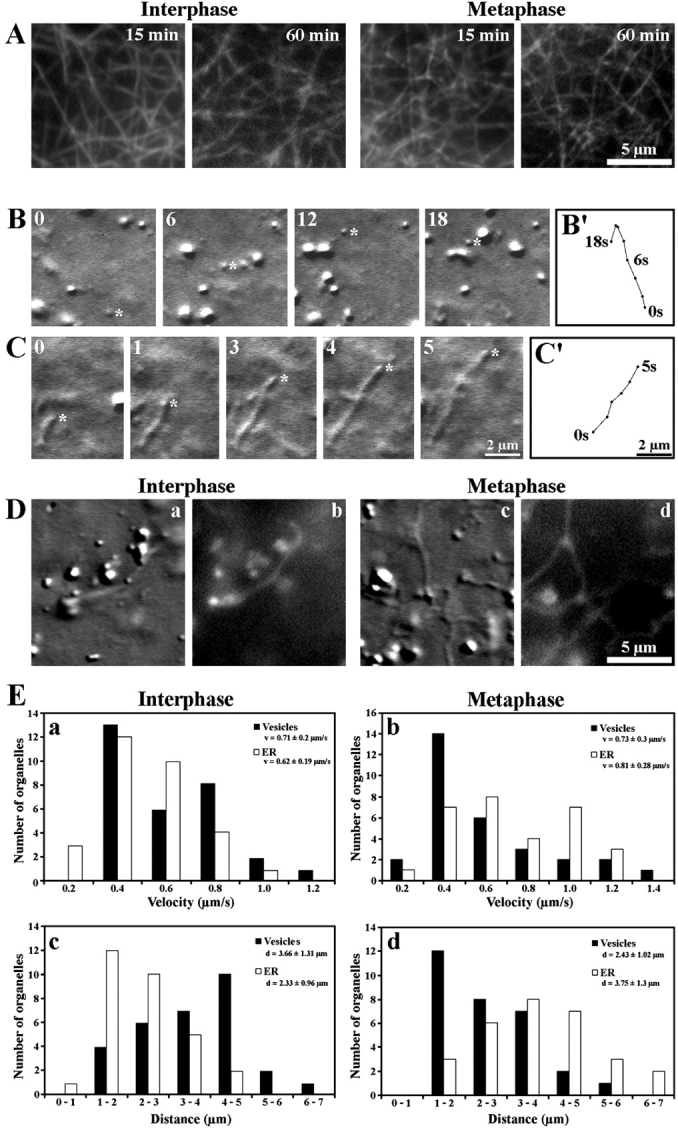
Reconstitution of organelle motility on F-actin in extracts isolated from Xenopus eggs. (A) Stability of an F-actin network in I (Interphase) and M (Metaphase) extracts visualized by fluorescence microscopy after 15 and 60 min of incubation with 0.5 μM rhodamine-phalloidin. (B) Movement of a globular vesicle on F-actin monitored by DIC microscopy. Selected frames of a video sequence covering 18 s are shown. (B′) Track diagram mapping the movement of the vesicle marked by an asterisk in B. (C) Movement of a tubular organelle (asterisk) on F-actin monitored by DIC microscopy. Selected frames of a video sequence covering 5 s are shown. (C′) Track diagram mapping the movement of the tubular organelle shown in C. The numbers in the top left corner of B and C indicate the time points of image acquisition. (D) Tubular organelles in I (Interphase) and M (Metaphase) extracts visualized by DIC microscopy (a and c) and by fluorescence microscopy after ER staining with DiOC6 (b and d). (E) Motion analysis of organelle movement on F-actin. (a and b) Distribution of the velocities and average velocities (v) attained by 30 globular vesicles (Vesicles) in I extracts (a) and by 30 tubular organelles (ER) in meiosis II M extracts (b). (c and d) Distributions of the frequencies of run distances and average run distance (d) attained by 30 globular vesicles (Vesicles) in I extracts (c) and by 30 tubular organelles (ER) in meiosis II M extracts (d). See also videos 1 and 2 available at http://www.jcb.org/cgi/content/full/jcb.200204065/DC1
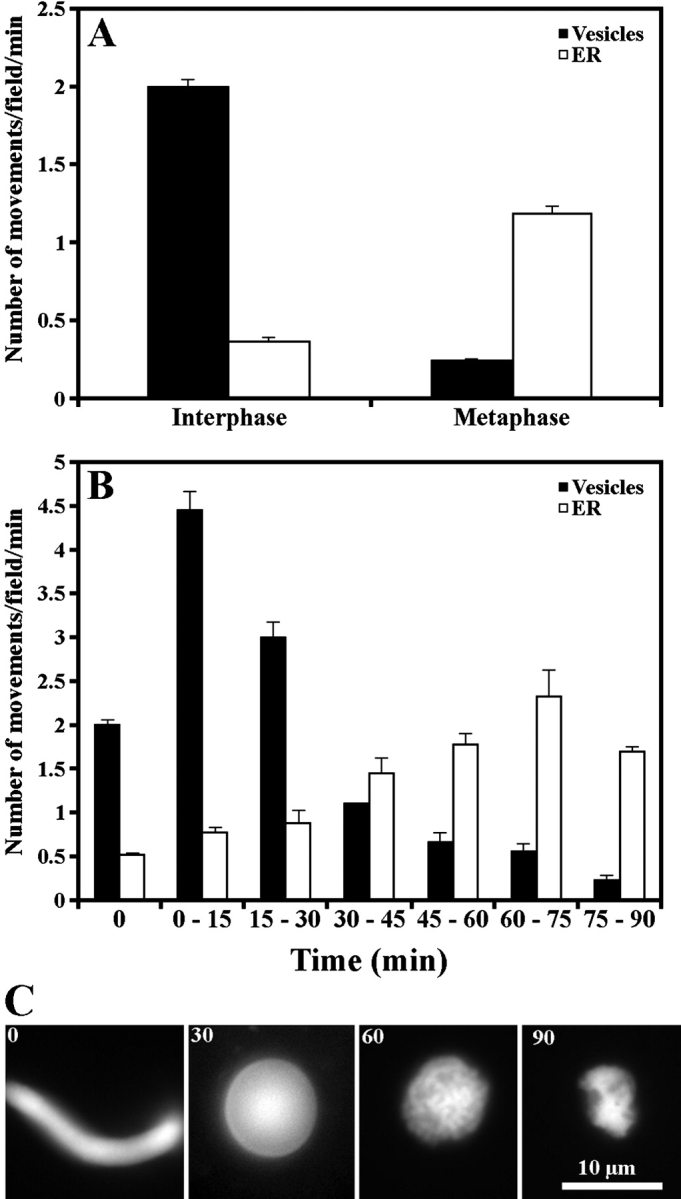
Quantitative analysis of actin-dependent organelle movement. (A) Motile activities of globular vesicles (Vesicles) and tubular organelles (ER) in I and meiosis II M extracts. (B) Motile activities of globular vesicles (Vesicles) and tubular organelles (ER) during transition from interphase to mitotic metaphase induced by CYCΔ. The values at the 0-min time point indicate the motile activity in the same I extract in the absence of CYCΔ. (C) Determination of cell cycle status during transition from interphase to mitotic metaphase induced by CYCΔ using morphology assay of added sperm nuclei.

F-actin–dependent ER network formation in M extracts. (A) An extensive network of ER was visualized with both DIC microscopy (a) and fluorescence microscopy after staining with DiOC6 (b) and immunofluorescence labeling with an antibody against ECaSt/PDI (e) in M extracts. No membrane network was detected in I extracts (c, d, and f). (B) An ER network was also observed in mitotic M extract obtained by incubation of I extracts with CYCΔ (a). An ER network did not form in the presence of 5 μM cytochalasin D (b). See also video 3 available at http://www.jcb.org/cgi/content/full/jcb.200204065/DC1 .
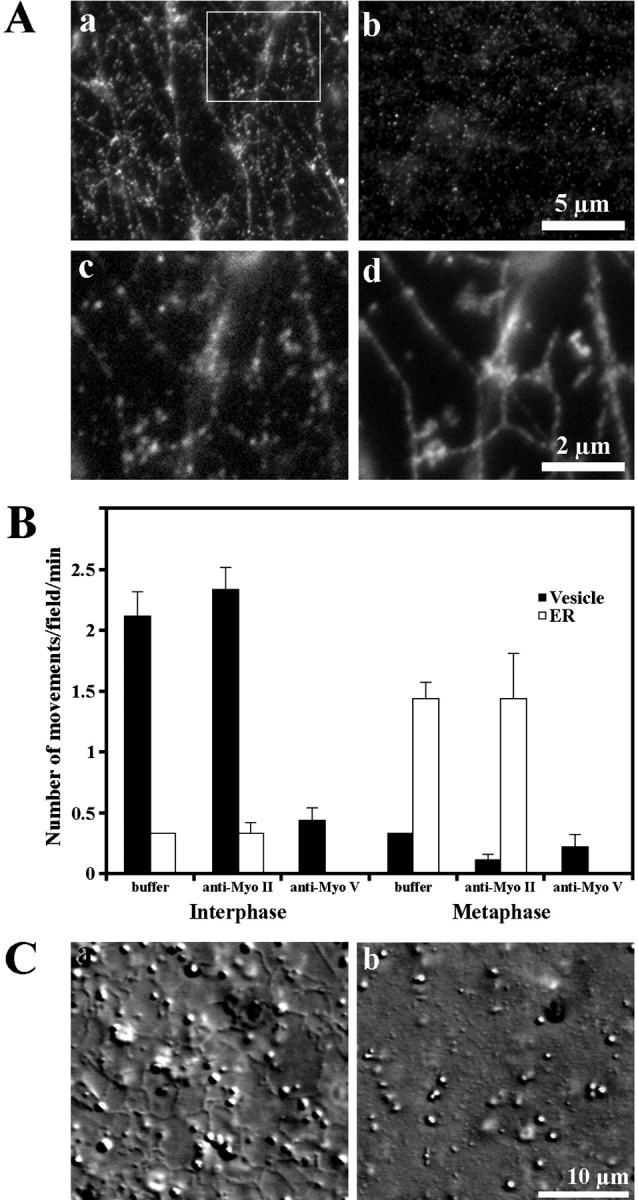
Identification of myosin V as a motor responsible for organelle motility and ER network formation. (A) The DIL2 antibody against myosin Va (a) but not a control antibody against myosin II (b) labeled the network in fixed preparations of M extract. The labeling of ER network with DIL2 antibody (c) and with an antibody against ECaSt/PDI (d) showed strong colocalization of myosin V immunosignal with ECaSt/PDI immunosignal. Panels c and d represent the region marked in panel a. (B) Inhibition of organelle motility on F-actin by an antibody against myosin Va. Motile activity in I and M extracts treated with DIL2 antibody (anti-Myo V) and a control antibody (anti-Myo II). (C) In M extracts ER network formation occurred in the presence of anti–myosin II (a) but not in the presence of DIL2 antibody (b).
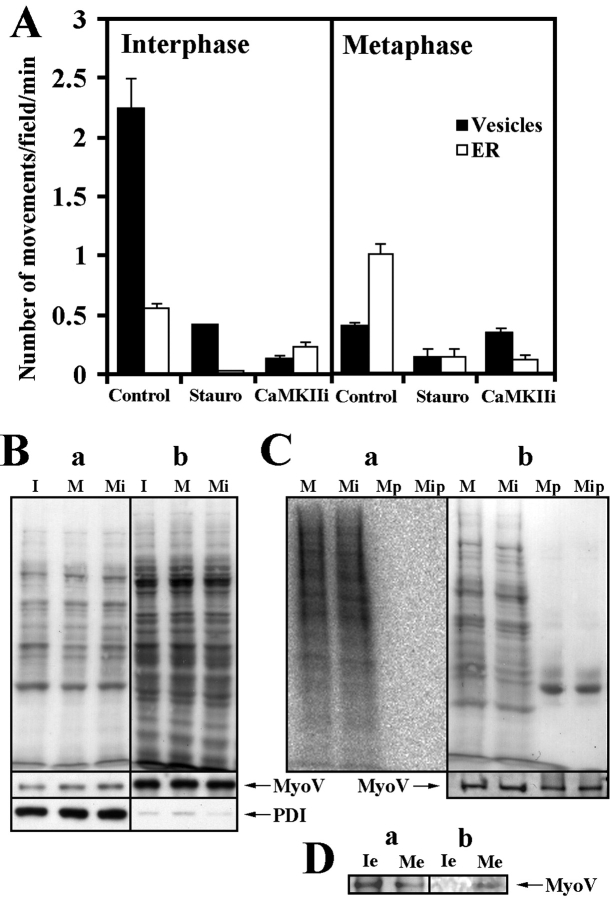
Effect of protein kinase activity on myosin V–based organelle motility. (A) 2 μM of staurosporine (Stauro) or 2 μM of CaMKII inhibitor (CaMKIIi) inhibited organelle motility in both I and meiosis II M extracts. The same effect was observed in mitotic M extract (data not depicted). (B) Immunoblotting of membrane fractions (a) and membrane free fractions (b) isolated from I extract (I), M extract (M), and M extract treated with CaMKII inhibitor (Mi) with DIL2 antibody (MyoV) and ER marker EcaSt/PDI (PDI). The equal amount of myosin V and EcaSt/PDI associated with membrane fractions. (Top) Ponceau S staining to demonstrate approximately equal protein load. (C) Phosphoimaging of membranes isolated from 32P-labeled mitotic M extract (M) and 32P-labeled mitotic M extract treated with CaMKII inhibitor (Mi) and myosin V immunoprecipitates (Mp and Mip) from the respective membrane fractions (a). (b) Ponceau S staining (top) and immunoblotting with DIL2 antibody (MyoV) (bottom) as in panel a to demonstrate approximately equal protein load and the presence of myosin V in membrane fractions and immunoprecipitates. (D) Immunoblotting (a) and phosphoimaging (b) of myosin V immnunoprecipitates from the nonfractionated I extract (Ie) and M extract (Me).
Similar articles
-
Cell cycle regulation of myosin-V by calcium/calmodulin-dependent protein kinase II.Science. 2001 Aug 17;293(5533):1317-20. doi: 10.1126/science.1061086. Science. 2001. PMID: 11509731
-
Regulation of melanosome movement in the cell cycle by reversible association with myosin V.J Cell Biol. 1999 Sep 20;146(6):1265-76. doi: 10.1083/jcb.146.6.1265. J Cell Biol. 1999. PMID: 10491390 Free PMC article.
-
Microinjected F-actin into dividing newt eggs moves toward the next cleavage furrow together with Ca2+ stores with inositol 1,4,5-trisphosphate receptor in a microtubule- and microtubule motor-dependent manner.Ital J Anat Embryol. 2008 Jul-Sep;113(3):143-51. Ital J Anat Embryol. 2008. PMID: 19205586
-
Dynamics and inheritance of the endoplasmic reticulum.J Cell Sci. 2004 Jun 15;117(Pt 14):2871-8. doi: 10.1242/jcs.01286. J Cell Sci. 2004. PMID: 15197242 Review.
-
Vesicle transport: the role of actin filaments and myosin motors.Microsc Res Tech. 1999 Oct 15;47(2):93-106. doi: 10.1002/(SICI)1097-0029(19991015)47:2<93::AID-JEMT2>3.0.CO;2-P. Microsc Res Tech. 1999. PMID: 10523788 Review.
Cited by
-
Endoplasmic reticulum remains continuous and undergoes sheet-to-tubule transformation during cell division in mammalian cells.J Cell Biol. 2007 Dec 3;179(5):895-909. doi: 10.1083/jcb.200705112. J Cell Biol. 2007. PMID: 18056408 Free PMC article.
-
A tubule-sheet continuum model for the mechanism of nuclear envelope assembly.Dev Cell. 2023 May 22;58(10):847-865.e10. doi: 10.1016/j.devcel.2023.04.003. Epub 2023 Apr 24. Dev Cell. 2023. PMID: 37098350 Free PMC article.
-
Intertwined and Finely Balanced: Endoplasmic Reticulum Morphology, Dynamics, Function, and Diseases.Cells. 2021 Sep 7;10(9):2341. doi: 10.3390/cells10092341. Cells. 2021. PMID: 34571990 Free PMC article. Review.
-
Myosin Va participates in acrosomal formation and nuclear morphogenesis during spermatogenesis of Chinese mitten crab Eriocheir sinensis.PLoS One. 2010 Sep 14;5(9):e12738. doi: 10.1371/journal.pone.0012738. PLoS One. 2010. PMID: 20856877 Free PMC article.
-
Fast anterograde transport of herpes simplex virus: role for the amyloid precursor protein of alzheimer's disease.Aging Cell. 2003 Dec;2(6):305-18. doi: 10.1046/j.1474-9728.2003.00069.x. Aging Cell. 2003. PMID: 14677633 Free PMC article.
References
-
- Felix, M.A., P.R. Clarke, J. Verde, and E. Karsenti. 1993. Xenopus egg extracts as a system for studing mitosis. The Cell Cycle—A Practical Approach. P. Fantes and R. Brooks, editors. Oxford University Press Inc., New York. 253–283.
-
- Glotzer, M., A.W. Murray, and M.W. Kirschner. 1991. Cyclin is degraded by the ubiquitin pathway. Nature. 349:132–138. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
