

The Xenopus Xmus101 protein is required for the recruitment of Cdc45 to origins of DNA replication
- PMID: 12438414
- PMCID: PMC2173091
- DOI: 10.1083/jcb.200207090
The Xenopus Xmus101 protein is required for the recruitment of Cdc45 to origins of DNA replication
Abstract
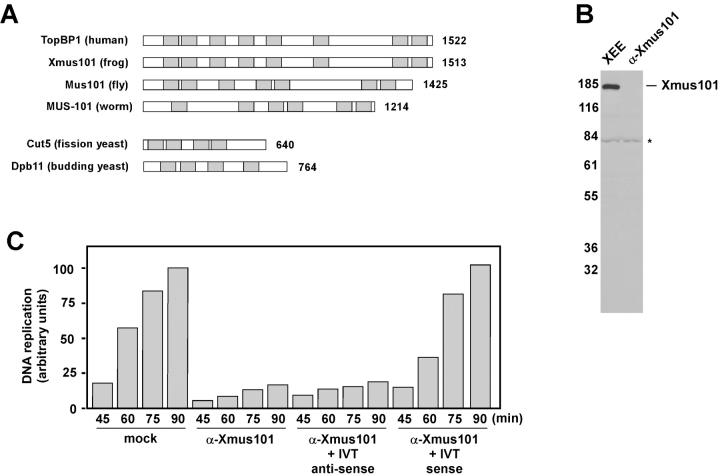
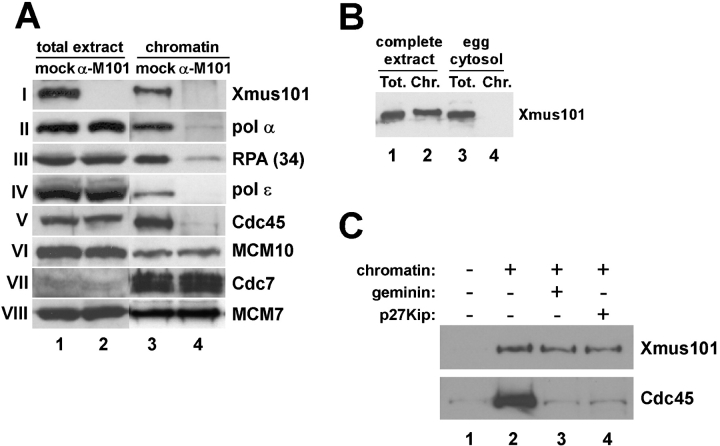
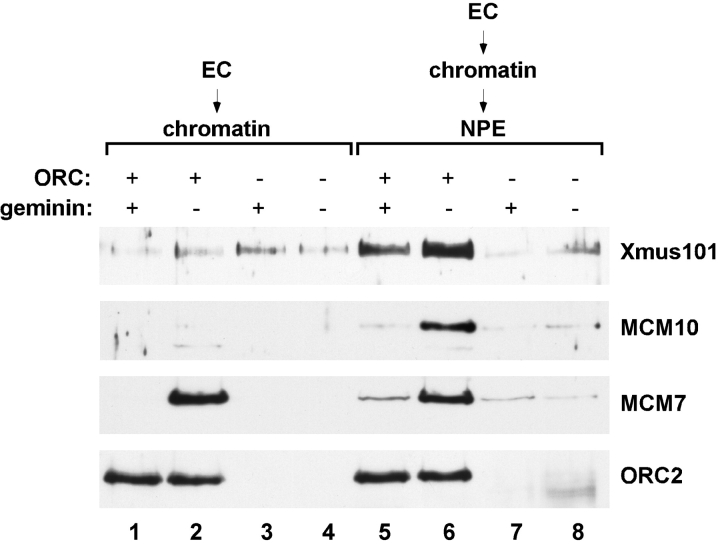
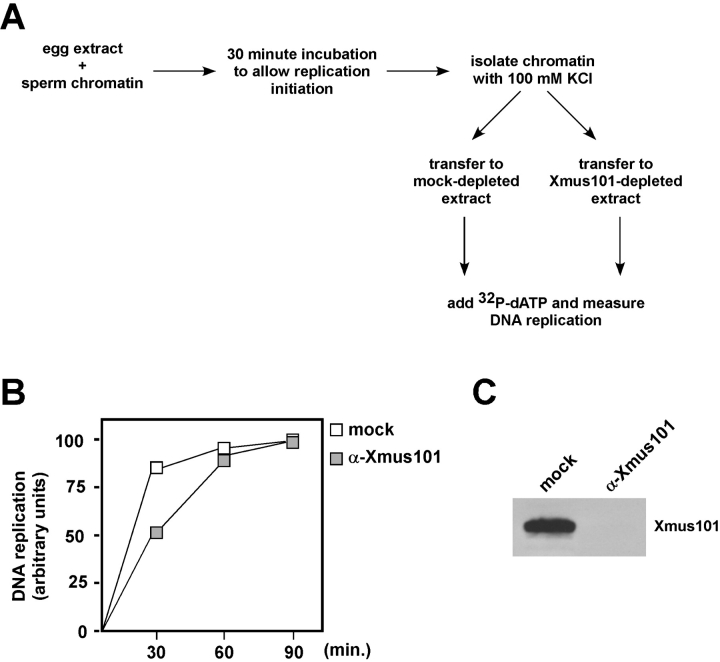
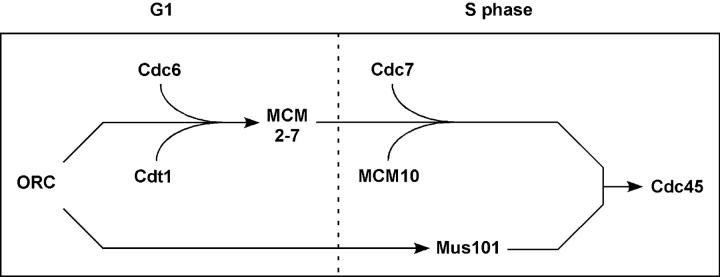
The initiation of eukaryotic DNA replication involves origin recruitment and activation of the MCM2-7 complex, the putative replicative helicase. Mini-chromosome maintenance (MCM)2-7 recruitment to origins in G1 requires origin recognition complex (ORC), Cdt1, and Cdc6, and activation at G1/S requires MCM10 and the protein kinases Cdc7 and S-Cdk, which together recruit Cdc45, a putative MCM2-7 cofactor required for origin unwinding. Here, we show that the Xenopus BRCA1 COOH terminus repeat-containing Xmus101 protein is required for loading of Cdc45 onto the origin. Xmus101 chromatin association is dependent on ORC, and independent of S-Cdk and MCM2-7. These results define a new factor that is required for Cdc45 loading. Additionally, these findings indicate that the initiation complex assembly pathway bifurcates early, after ORC association with the origin, and that two parallel pathways, one controlled by MCM2-7, and the other by Xmus101, cooperate to load Cdc45 onto the origin.
Figures
Similar articles
-
MCM2-7 complexes bind chromatin in a distributed pattern surrounding the origin recognition complex in Xenopus egg extracts.J Biol Chem. 2002 Sep 6;277(36):33049-57. doi: 10.1074/jbc.M204438200. Epub 2002 Jun 26. J Biol Chem. 2002. PMID: 12087101
-
A requirement for MCM7 and Cdc45 in chromosome unwinding during eukaryotic DNA replication.EMBO J. 2004 Sep 15;23(18):3667-76. doi: 10.1038/sj.emboj.7600369. Epub 2004 Aug 26. EMBO J. 2004. PMID: 15329670 Free PMC article.
-
Xenopus Mcm10 binds to origins of DNA replication after Mcm2-7 and stimulates origin binding of Cdc45.Mol Cell. 2002 Feb;9(2):233-40. doi: 10.1016/s1097-2765(02)00456-2. Mol Cell. 2002. PMID: 11864598
-
Behavior of replication origins in Eukaryota - spatio-temporal dynamics of licensing and firing.Cell Cycle. 2015;14(14):2251-64. doi: 10.1080/15384101.2015.1056421. Epub 2015 Jun 1. Cell Cycle. 2015. PMID: 26030591 Free PMC article. Review.
-
Control of DNA replication licensing in a cell cycle.Genes Cells. 2002 Jun;7(6):523-34. doi: 10.1046/j.1365-2443.2002.00544.x. Genes Cells. 2002. PMID: 12059957 Review.
Cited by
-
Role for Rif1 in the checkpoint response to damaged DNA in Xenopus egg extracts.Cell Cycle. 2012 Mar 15;11(6):1183-94. doi: 10.4161/cc.11.6.19636. Epub 2012 Mar 15. Cell Cycle. 2012. PMID: 22391207 Free PMC article.
-
The Mre11-Rad50-Nbs1 (MRN) complex has a specific role in the activation of Chk1 in response to stalled replication forks.Mol Biol Cell. 2013 May;24(9):1343-53. doi: 10.1091/mbc.E13-01-0025. Epub 2013 Mar 6. Mol Biol Cell. 2013. PMID: 23468519 Free PMC article.
-
BRCA1 and its toolbox for the maintenance of genome integrity.Nat Rev Mol Cell Biol. 2010 Feb;11(2):138-48. doi: 10.1038/nrm2831. Epub 2009 Dec 23. Nat Rev Mol Cell Biol. 2010. PMID: 20029420 Free PMC article. Review.
-
Differential binding of replication proteins across the human c-myc replicator.Mol Cell Biol. 2006 Jul;26(14):5270-83. doi: 10.1128/MCB.02137-05. Mol Cell Biol. 2006. PMID: 16809765 Free PMC article.
-
The DNA unwinding element binding protein DUE-B interacts with Cdc45 in preinitiation complex formation.Mol Cell Biol. 2010 Mar;30(6):1495-507. doi: 10.1128/MCB.00710-09. Epub 2010 Jan 11. Mol Cell Biol. 2010. PMID: 20065034 Free PMC article.
References
-
- Bell, S.P., and A. Dutta. 2002. DNA replication in eukaryotic cells. Annu. Rev. Biochem. 71:333–374. - PubMed
-
- Edwards, M.C., A.V. Tutter, C. Cvetic, C.H. Gilbert, T.A. Prokhorova, and J.C. Walter. 2002. MCM2-7 complexes bind chromatin in a distributed pattern surrounding the origin recognition complex in Xenopus egg extracts. J. Biol. Chem. 277:33049–33057. - PubMed
-
- Harlow, E., and D. Lane. 1988. Antibodies, a Laboratory Manual. Cold Spring Harbor Press, Cold Spring Harbor, NY. 312 pp.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
