

African swine fever virus polyproteins pp220 and pp62 assemble into the core shell
- PMID: 12438573
- PMCID: PMC136718
- DOI: 10.1128/jvi.76.24.12473-12482.2002
African swine fever virus polyproteins pp220 and pp62 assemble into the core shell
Abstract
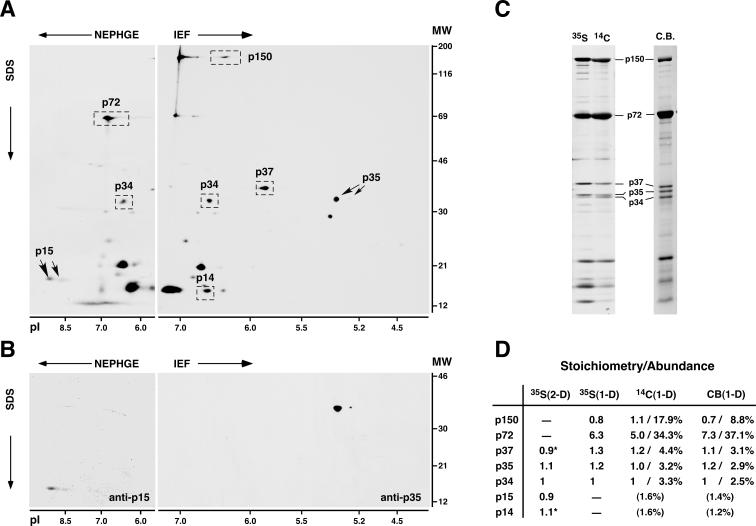
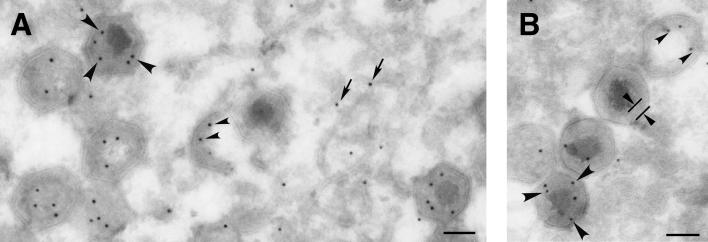
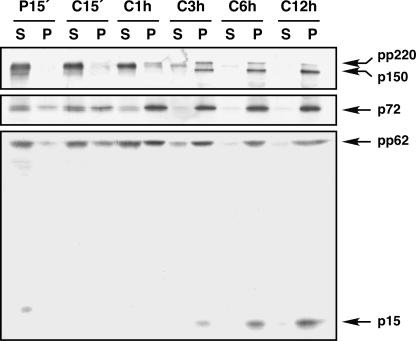
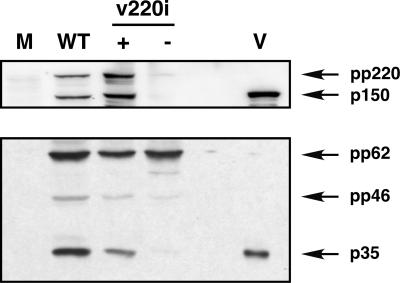
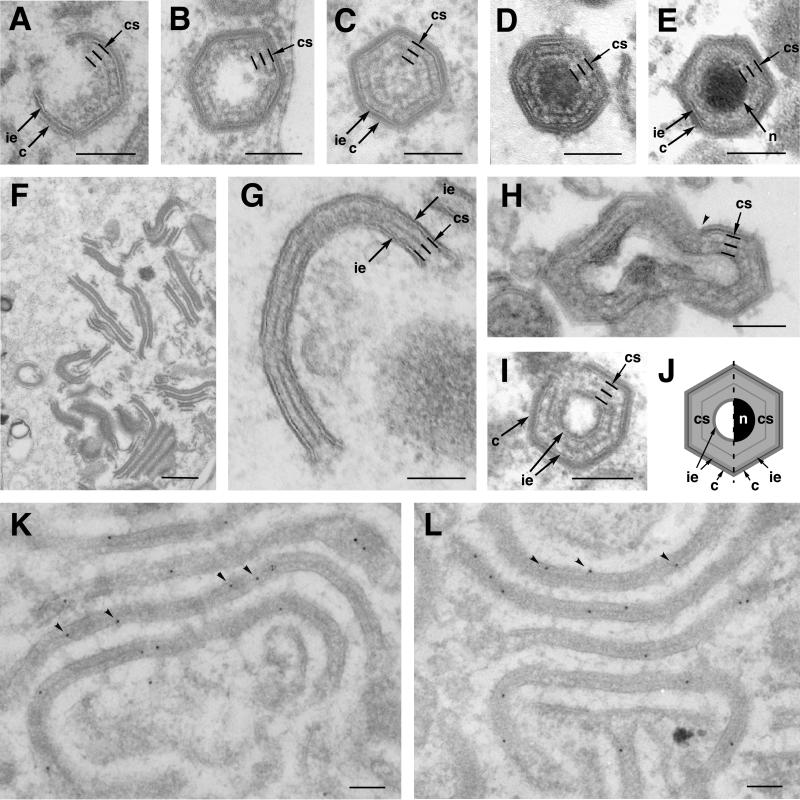
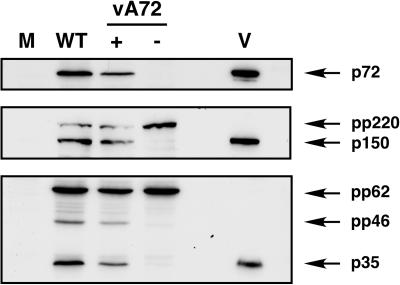
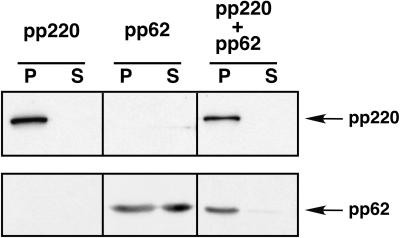
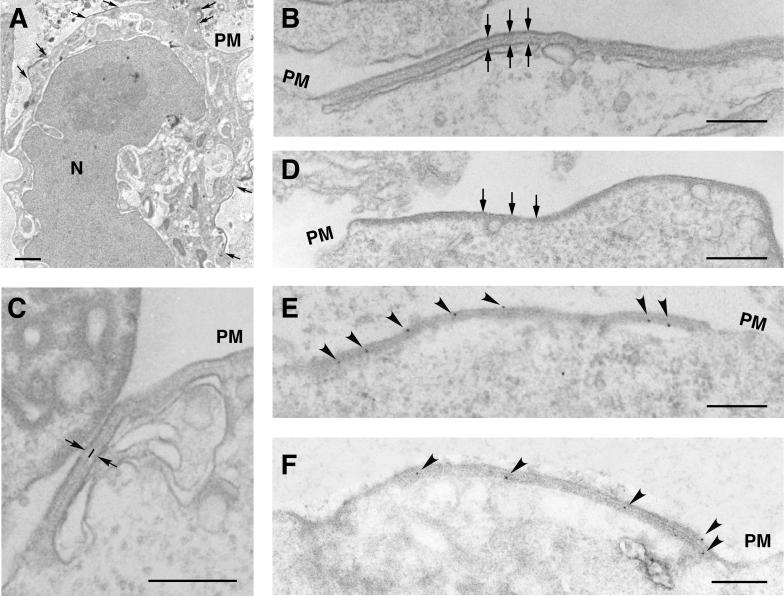
African swine fever virus (ASFV), a complex enveloped DNA virus, expresses two polyprotein precursors, pp220 and pp62, which after proteolytic processing give rise to several major components of the virus particle. We have analyzed the structural role of polyprotein pp62, the precursor form of mature products p35 and p15, in virus morphogenesis. Densitometric analysis of one- and two-dimensional gels of purified virions showed that proteins p35 and p15, as well as the pp220-derived products, are present in equimolecular amounts in the virus particle. Immunoelectron microscopy revealed that the pp62-derived products localize at the core shell, a matrix-like domain placed between the DNA-containing nucleoid and the inner envelope, where the pp220-derived products are also localized. Pulse-chase experiments indicated that the processing of both polyprotein precursors is concomitant with virus assembly. Furthermore, using inducible ASFV recombinants, we show that pp62 processing requires the expression of the pp220 core precursor, whereas the processing of both precursors pp220 and pp62 is dependent on expression of the major capsid protein p72. Interestingly, when p72 expression is blocked, unprocessed pp220 and pp62 polyproteins assemble into aberrant zipper-like elements consisting of an elongated membrane-bound protein structure reminiscent of the core shell. Moreover, the two polyproteins, when coexpressed in COS cells, interact with each other to form zipper-like structures. Together, these findings indicate that the mature products derived from both polyproteins, which collectively account for about 30% of the virion protein mass, are the basic components of the core shell and that polyprotein processing represents a maturational process related to ASFV morphogenesis.
Figures
References
-
- Alonso, C., J. Miskin, B. Hernáez, P. Fernández-Zapatero, L. Soto, C. Cantó, I. Rodríguez-Crespo, L. Dixon, and J. M. Escribano. 2001. African swine fever virus protein p54 interacts with the microtubular motor complex through direct binding to light-chain dynein. J. Virol. 75:9819-9827. - PMC - PubMed
-
- Alves de Matos, A. P., and Z. G. Carvalho. 1993. African swine fever virus interaction with microtubules. Biol. Cell 78:229-234. - PubMed
-
- Andrés, G., C. Simón-Mateo, and E. Viñuela. 1993. Characterization of two African swine fever virus 220-kDa proteins: a precursor of the major structural protein p150 and an oligomer of phosphoprotein p32. Virology 194:284-293. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
