

Transcriptional regulation of the K1 gene product of Kaposi's sarcoma-associated herpesvirus
- PMID: 12438583
- PMCID: PMC136681
- DOI: 10.1128/jvi.76.24.12574-12583.2002
Transcriptional regulation of the K1 gene product of Kaposi's sarcoma-associated herpesvirus
Abstract
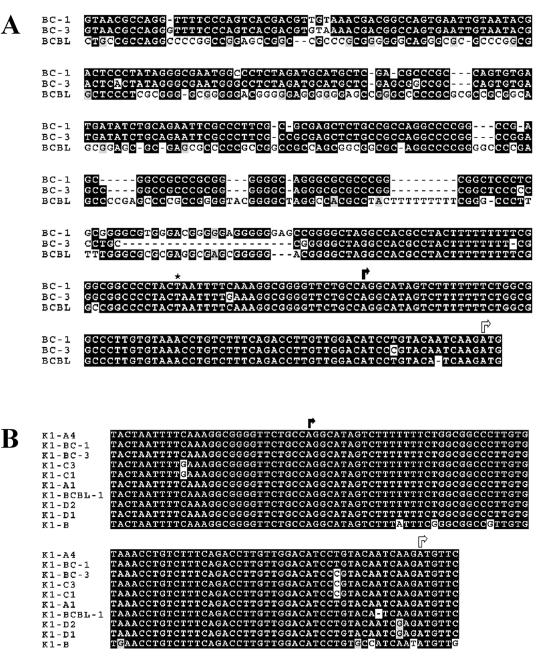
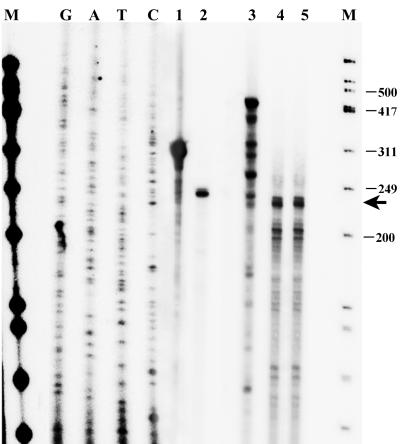
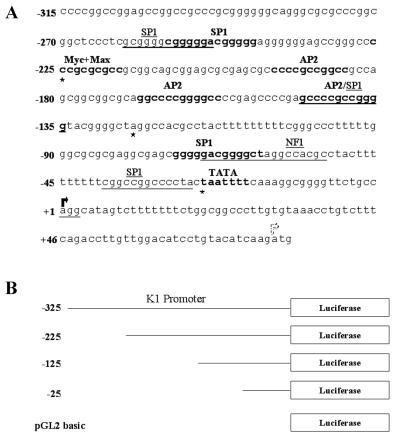
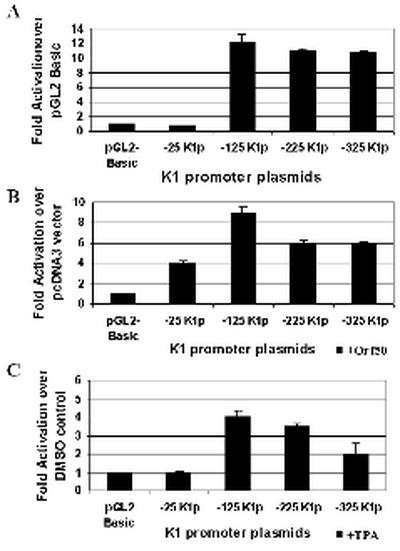
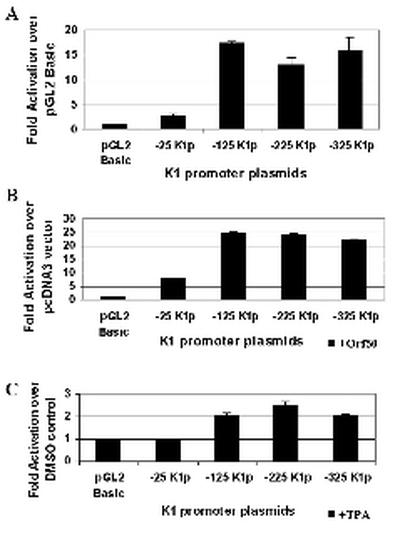
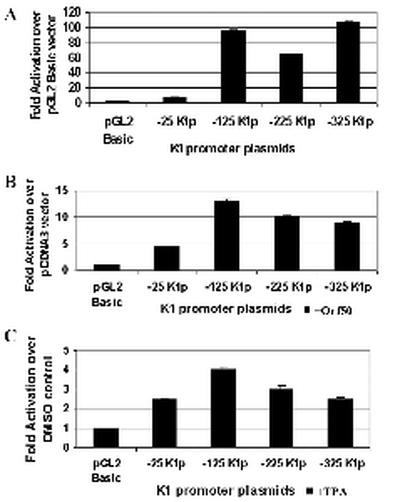
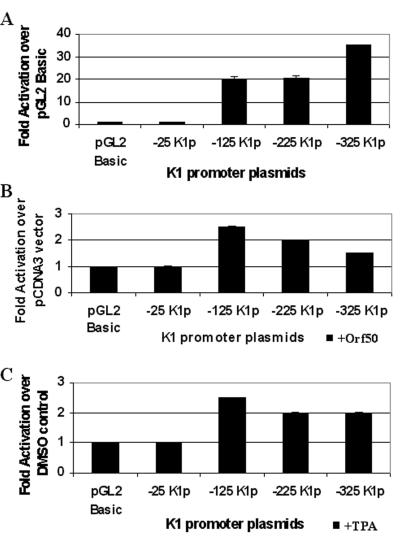
The K1 protein of Kaposi's sarcoma-associated herpesvirus (KSHV) has been shown to be a transforming protein capable of inducing morphological changes and focus formation in rodent fibroblasts. K1 can activate B-cell receptor (BCR) signaling and upregulate activity of the NFAT and NF-kappaB transcription factors. In order to understand the regulation of K1 gene expression, we have analyzed sequences upstream of the K1 gene to identify the K1 promoter element. We have performed 5' rapid amplification of cDNA ends as well as a nuclease protection assay to map the transcriptional start site of the KSHV K1 transcript. The K1 transcriptional start site lies 75 bp upstream of the translation start site. Sequences upstream of the K1 gene were characterized for their ability to activate a luciferase reporter gene in 293 epithelial cells, KSHV-negative B cells (BJAB), KSHV-positive B cells (BCBL-1), and KS tumor-derived endothelial cells (SLK-KS(-)). We found that a 125-bp sequence upstream of the K1 transcript start site was sufficient to fully activate the luciferase reporter gene in all cell types tested. In addition, the viral transcription factor KSHV Orf50/Rta was capable of further activating this promoter element in 293, BJAB, and BCBL-1 cells but not in SLK-KS(-) cells. Promoter constructs containing additional sequences upstream of the 125-bp element did not show further augmentation of transcription in the presence or absence of KSHV Orf50.
Figures
References
-
- Alexander, L., L. Denenkamp, A. Knapp, M. Auerbach, B. Damania, and R. C. Desrosiers. 2000. The primary sequence of rhesus monkey rhadinovirus isolate 26-95: sequence similarities to Kaposi's sarcoma-associated herpesvirus and rhesus monkey rhadinovirus isolate 17577. J. Virol. 74:3388-3398. - PMC - PubMed
-
- Angel, P., M. Imagawa, R. Chiu, B. Stein, R. J. Imbra, H. J. Rahmsdorf, C. Jonat, P. Herrlich, and M. Karin. 1987. Phorbol ester-inducible genes contain a common cis element recognized by a TPA-modulated trans-acting factor. Cell 49:729-739. - PubMed
-
- Antman, K., and Y. Chang. 2000. Kaposi's sarcoma. N. Engl. J. Med. 342:1027-1038. - PubMed
-
- Boshoff, C., T. F. Schulz, M. M. Kennedy, A. K. Graham, C. Fisher, A. Thomas, J. O. McGee, R. A. Weiss, and J. J. O'Leary. 1995. Kaposi's sarcoma-associated herpesvirus infects endothelial and spindle cells. Nat. Med. 1:1274-1278. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
