

Mitotic chromosomes are chromatin networks without a mechanically contiguous protein scaffold
- PMID: 12438695
- PMCID: PMC137727
- DOI: 10.1073/pnas.232442599
Mitotic chromosomes are chromatin networks without a mechanically contiguous protein scaffold
Abstract
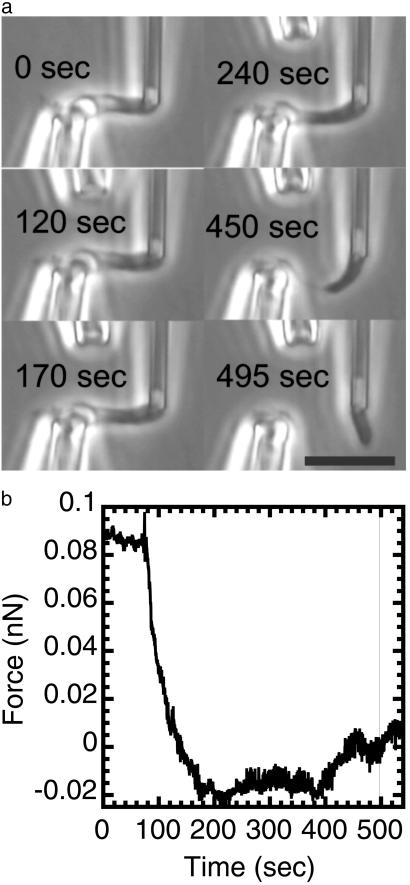
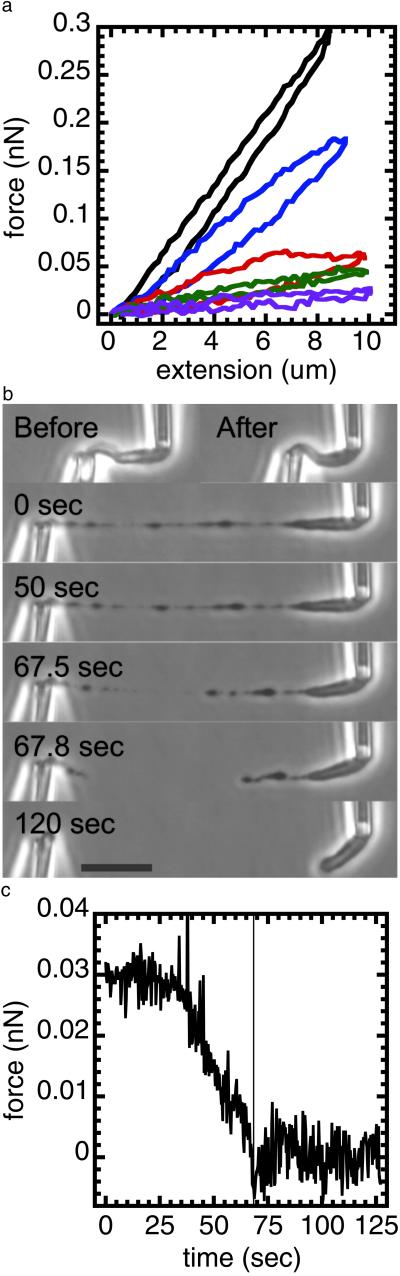
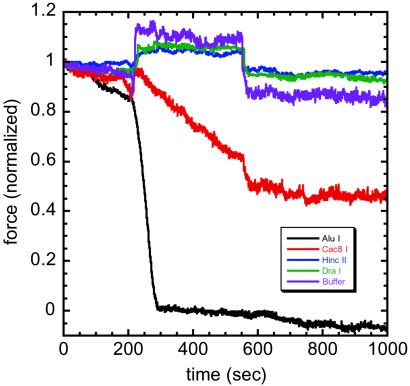
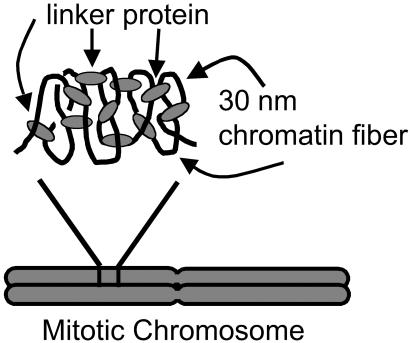
Isolated newt (Notophthalmus viridescens) chromosomes were studied by using micromechanical force measurement during nuclease digestion. Micrococcal nuclease and short-recognition-sequence blunt-cutting restriction enzymes first remove the native elastic response of, and then to go on to completely disintegrate, single metaphase newt chromosomes. These experiments rule out the possibility that the mitotic chromosome is based on a mechanically contiguous internal non-DNA (e.g., protein) "scaffold"; instead, the mechanical integrity of the metaphase chromosome is due to chromatin itself. Blunt-cutting restriction enzymes with longer recognition sequences only partially disassemble mitotic chromosomes and indicate that chromatin in metaphase chromosomes is constrained by isolated chromatin-crosslinking elements spaced by approximately 15 kb.
Figures
Comment in
-
Mitotic chromosome scaffold structure: new approaches to an old controversy.Proc Natl Acad Sci U S A. 2002 Dec 10;99(25):15855-7. doi: 10.1073/pnas.262672799. Epub 2002 Dec 2. Proc Natl Acad Sci U S A. 2002. PMID: 12461163 Free PMC article. No abstract available.
-
A silent revolution in chromosome biology.Nat Rev Mol Cell Biol. 2014 Jul;15(7):431. doi: 10.1038/nrm3817. Epub 2014 May 29. Nat Rev Mol Cell Biol. 2014. PMID: 24871800 No abstract available.
References
-
- Hirano T. (2000) Annu. Rev. Biochem. 69 115-144. - PubMed
-
- Dietzel S. & Belmont, A. S. (2001) Nat. Cell Biol. 3 767-770. - PubMed
-
- Paulson J. R. & Laemmli, U. K. (1977) Cell 12 817-828. - PubMed
-
- Marsden M. P. F. & Laemmli, U. K. (1979) Cell 17 849-858. - PubMed
-
- Laemmli U. K., Cheng, S. M., Adolph, K. W., Paulson, J. R., Brown, J. A. & Baumbach, W. R. (1978) Cold Spring Harbor Symp. Quant. Biol. 42 351-360. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
