

Immunopurified small nucleolar ribonucleoprotein particles pseudouridylate rRNA independently of their association with phosphorylated Nopp140
- PMID: 12446766
- PMCID: PMC139890
- DOI: 10.1128/MCB.22.24.8457-8466.2002
Immunopurified small nucleolar ribonucleoprotein particles pseudouridylate rRNA independently of their association with phosphorylated Nopp140
Abstract
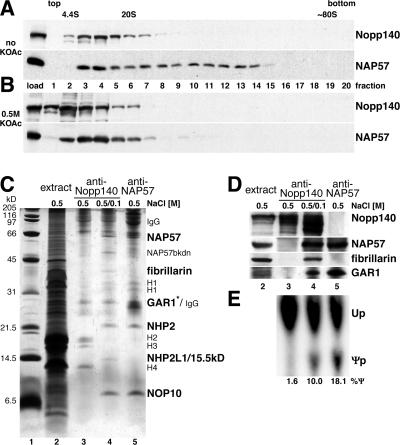
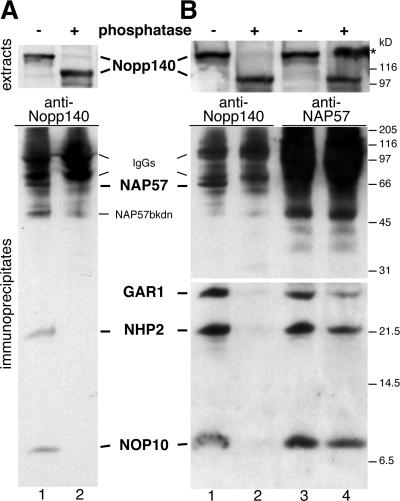
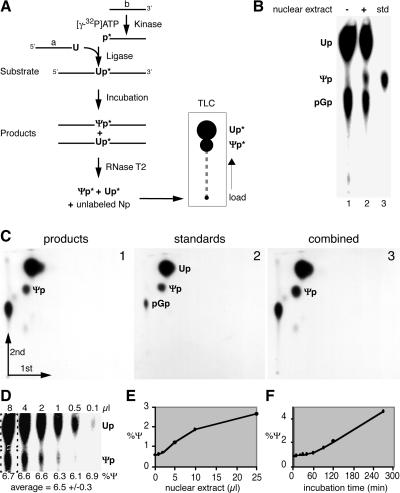
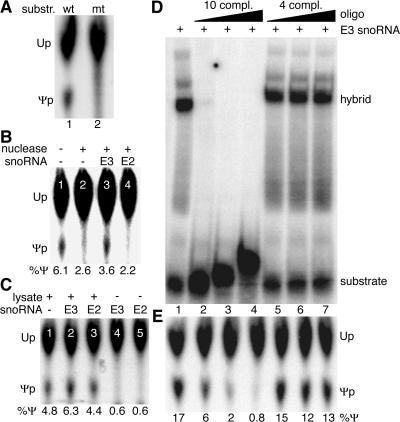
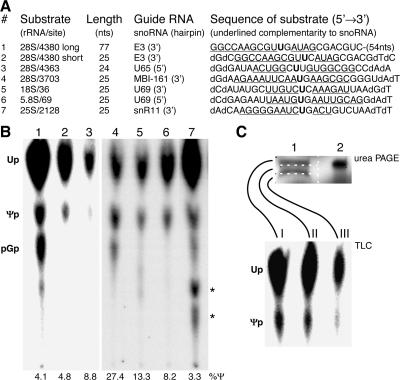
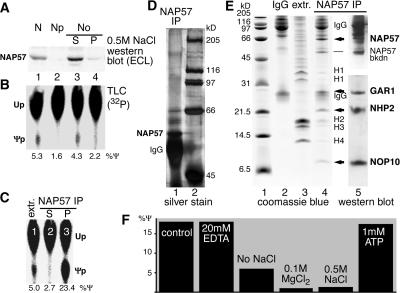
The isomerization of up to 100 uridines to pseudouridines (Psis) in eukaryotic rRNA is guided by a similar number of box H/ACA small nucleolar RNAs (snoRNAs), each forming a unique small nucleolar ribonucleoprotein particle (snoRNP) with the same four core proteins, NAP57 (also known as dyskerin or Cbf5p), GAR1, NHP2, and NOP10. Additionally, the nucleolar and Cajal body protein Nopp140 (Srp40p) associates with the snoRNPs. To understand the role of these factors in pseudouridylation, we established an in vitro assay system. Short site-specifically (32)P-labeled rRNA substrates were incubated with subcellular fractions, and the conversion of uridine to Psi was monitored by thin-layer chromatography after digestion to single nucleotides. Immunopurified box H/ACA core particles were sufficient for the reaction. SnoRNPs associated quantitatively and reversibly with Nopp140. However, pseudouridylation activity was independent of Nopp140, consistent with a chaperoning role for this highly phosphorylated protein. Although up to 14 bp between the snoRNA and rRNA were required for the in vitro reaction, rRNA pseudouridylation and release occurred in the absence of ATP and magnesium. These data suggest that substrate release takes place without RNA helicase activity but may be aided by the snoRNP core proteins.
Figures
References
-
- Bakin, A., and J. Ofengand. 1993. Four newly located pseudouridylate residues in Escherichia coli 23S ribosomal RNA are all at the peptidyltransferase center: analysis by the application of a new sequencing technique. Biochemistry 32:9754-9762. - PubMed
-
- Balakin, A. G., L. Smith, and M. J. Fournier. 1996. The RNA world of the nucleolus: two major families of small RNAs defined by different box elements with related functions. Cell 86:823-834. - PubMed
-
- Blobel, G., and V. Potter. 1966. Nuclei from rat liver: isolation method that combines purity with yield. Science 154:1662-1665. - PubMed
-
- Chang, M. S., H. Sasaki, M. S. Campbell, S. K. Kraeft, R. Sutherland, C. Y. Yang, Y. Liu, D. Auclair, L. Hao, H. Sonoda, L. H. Ferland, and L. B. Chen. 1999. HRad17 colocalizes with NHP2L1 in the nucleolus and redistributes after UV irradiation. J. Biol. Chem. 274:36544-36549. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases