

DRPEER: a motif in the extracellular vestibule conferring high Ca2+ flux rates in NMDA receptor channels
- PMID: 12451122
- PMCID: PMC6758750
- DOI: 10.1523/JNEUROSCI.22-23-10209.2002
DRPEER: a motif in the extracellular vestibule conferring high Ca2+ flux rates in NMDA receptor channels
Abstract
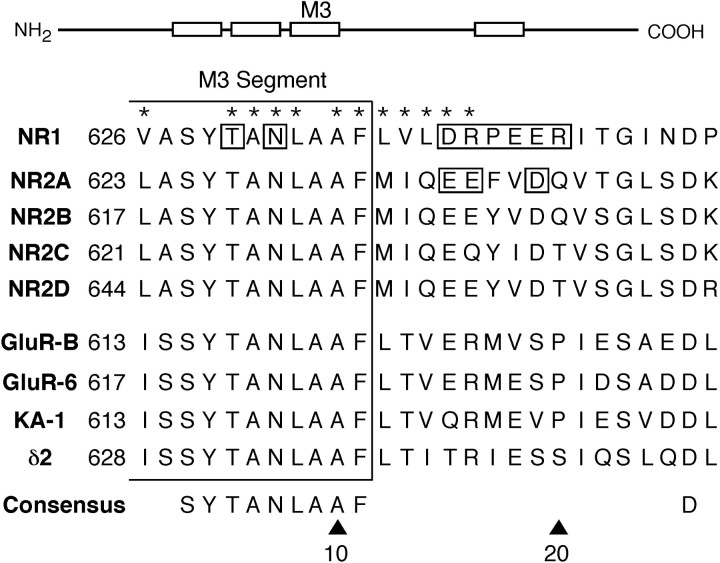
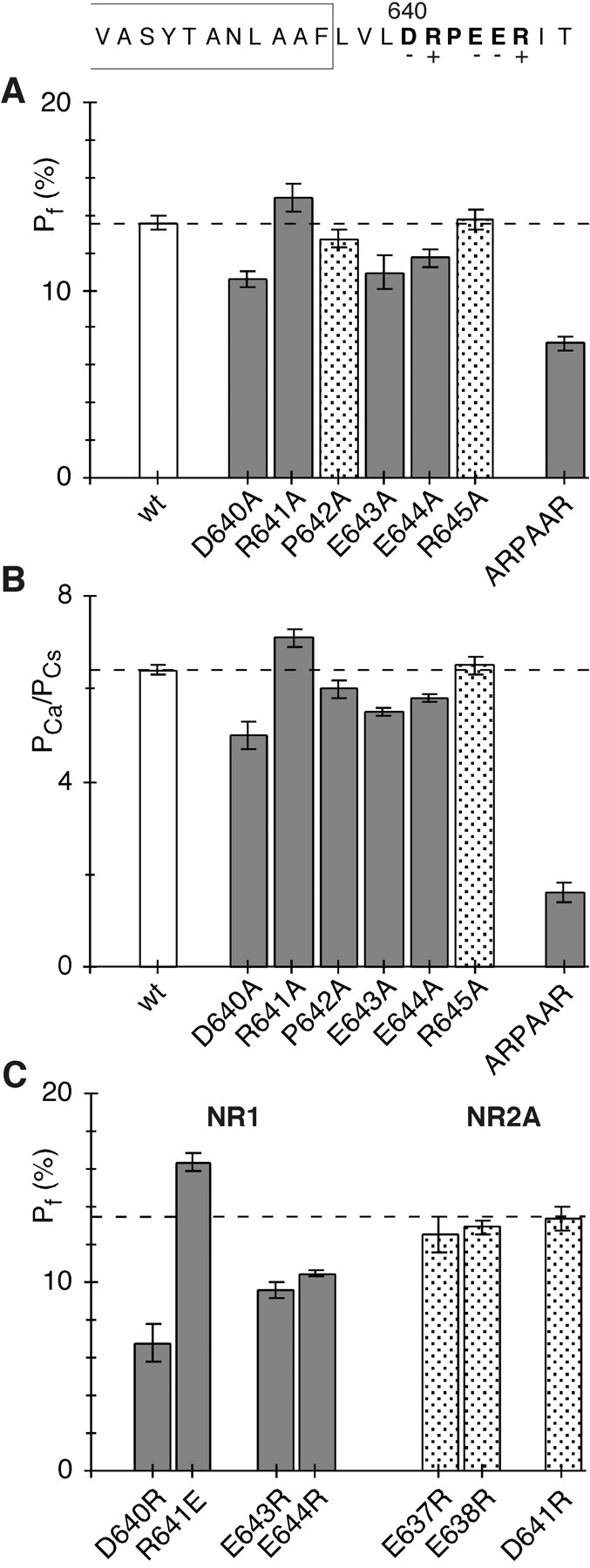
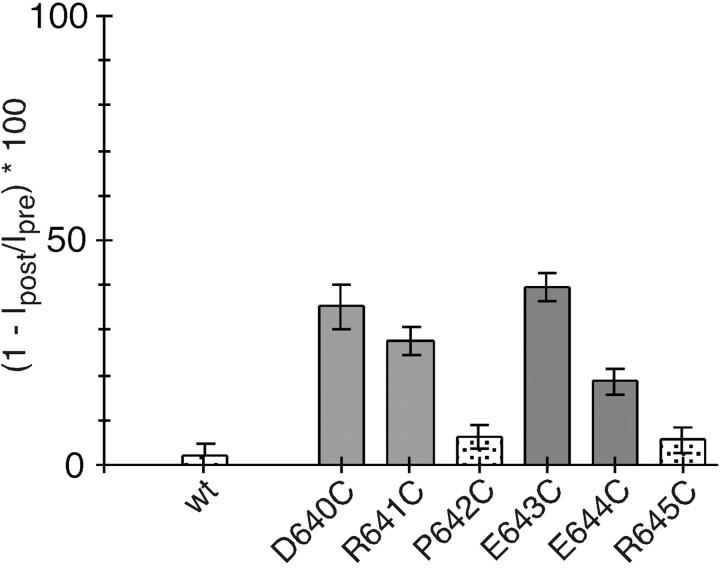
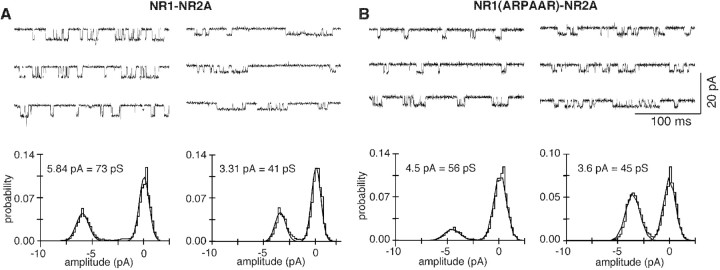
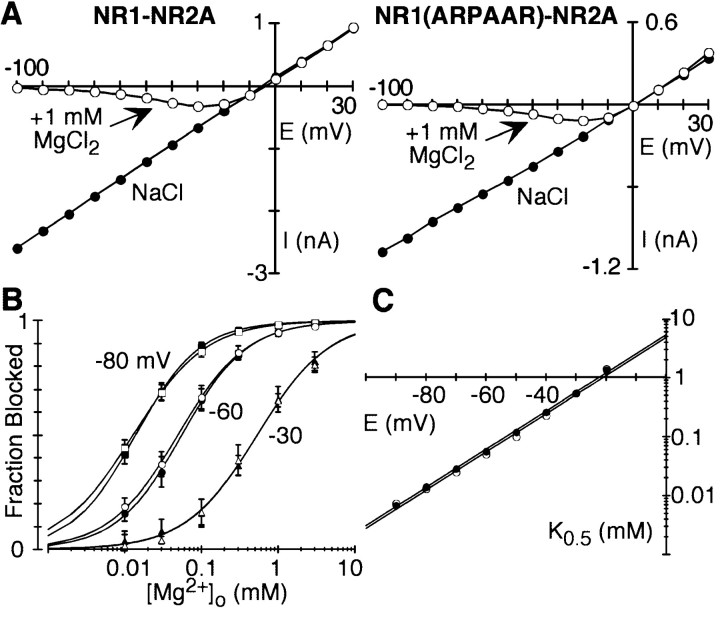
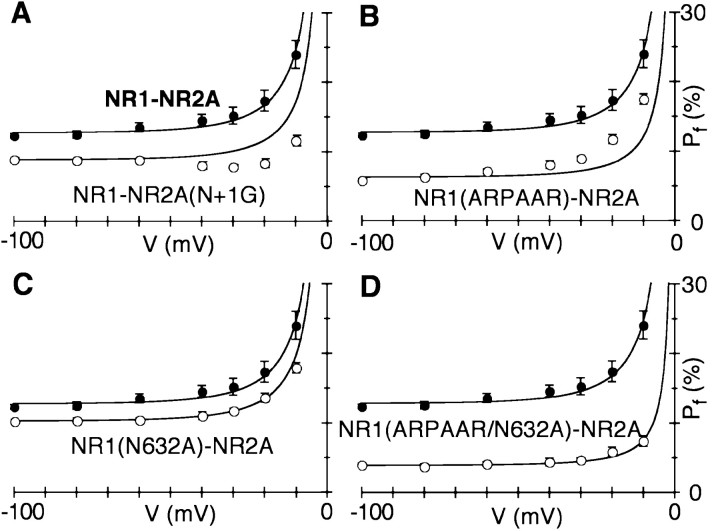
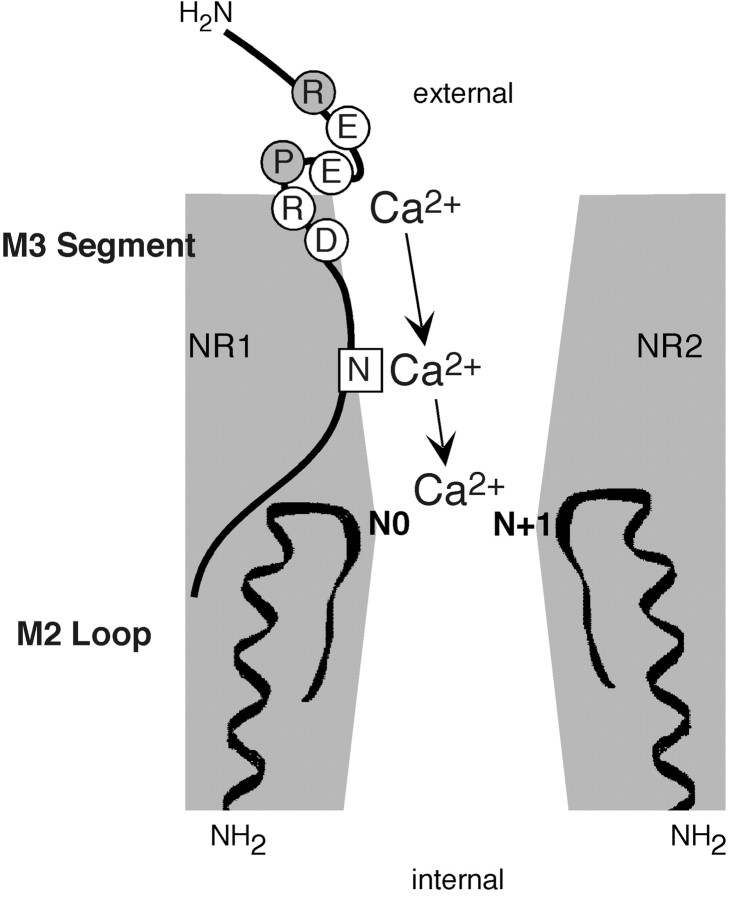
The high flux rate of Ca2+ through NMDA receptor (NMDAR) channels is critical for their biological function and may depend on a Ca2+ binding site in the extracellular vestibule. We screened substitutions of hydrophilic residues exposed in the vestibule and identified a cluster of charged residues and a proline, the DRPEER motif, positioned C terminal to M3, that is unique to the NR1 subunit. Charge neutralization or conversion of residues in DRPEER altered fractional Ca2+ currents in a manner consistent with its forming a binding site for Ca2+. Similarly, in a mutant channel in which all of the negative charges are neutralized (ARPAAR), the block by extracellular Ca2+ of single-channel current amplitudes is attenuated. In these same channels, the block by extracellular Mg2+ is unaffected. DRPEER is located extracellularly, and its contribution to Ca2+ influx is distinct from that of the narrow constriction. We conclude that key residues in DRPEER, acting as an external binding site for Ca2+, along with a conserved asparagine in the M3 segment proper, contribute to the high fractional Ca2+ currents in these channels under physiological conditions. Therefore, these domains represent critical molecular determinants of NMDAR function in synaptic physiology.
Figures
References
-
- Beck C, Wollmuth LP, Seeburg PH, Sakmann B, Kuner T. NMDAR channel segments forming the extracellular vestibule inferred from the accessibility of substituted cysteines. Neuron. 1999;22:559–570. - PubMed
-
- Burnashev N. Calcium permeability of glutamate-gated channels in the central nervous system. Curr Opin Neurobiol. 1996;6:311–317. - PubMed
-
- Burnashev N, Schoepfer R, Monyer H, Ruppersberg JP, Günther W, Seeburg PH, Sakmann B. Control by asparagine residues of calcium permeability and magnesium blockade in the NMDA receptor. Science. 1992;257:1415–1419. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous