

Cellular mechanisms regulating activity-dependent release of native brain-derived neurotrophic factor from hippocampal neurons
- PMID: 12451139
- PMCID: PMC6758764
- DOI: 10.1523/JNEUROSCI.22-23-10399.2002
Cellular mechanisms regulating activity-dependent release of native brain-derived neurotrophic factor from hippocampal neurons
Abstract
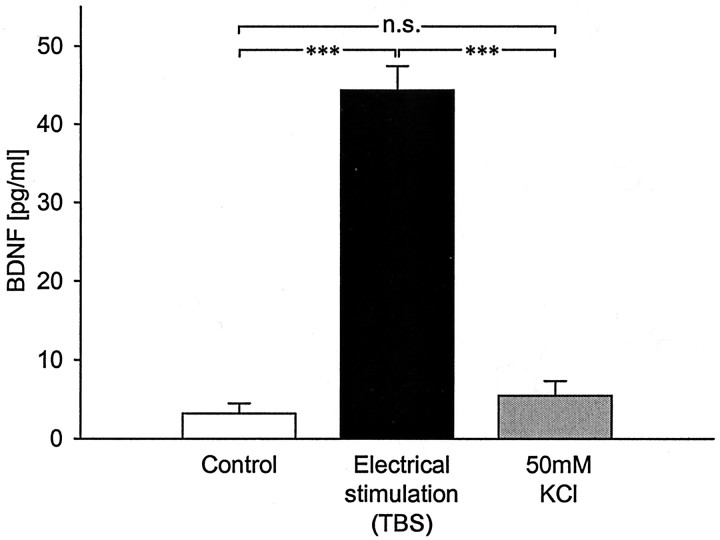
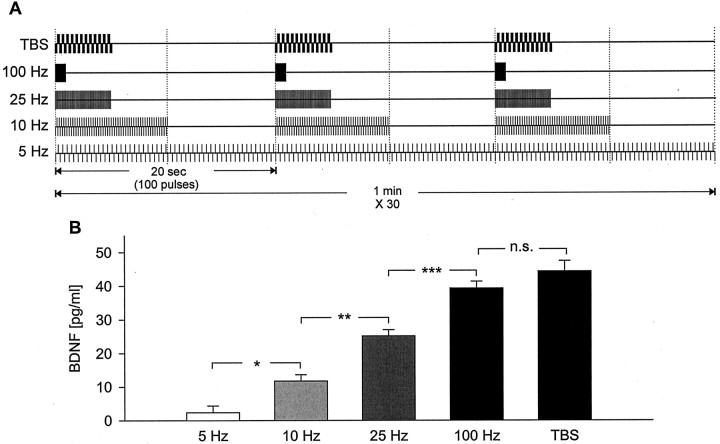
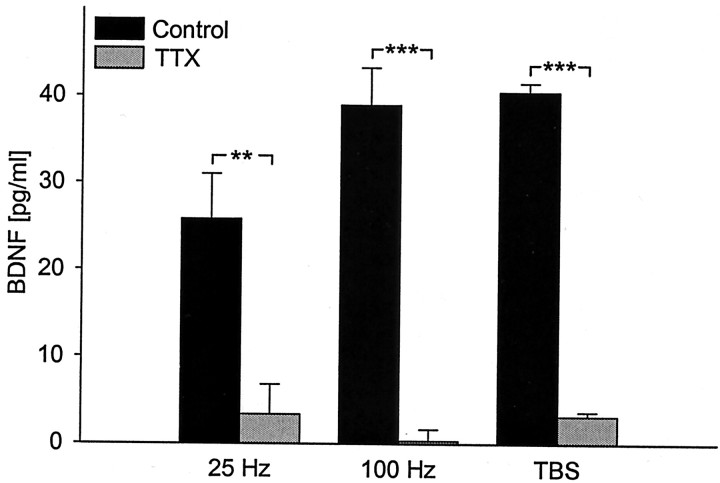
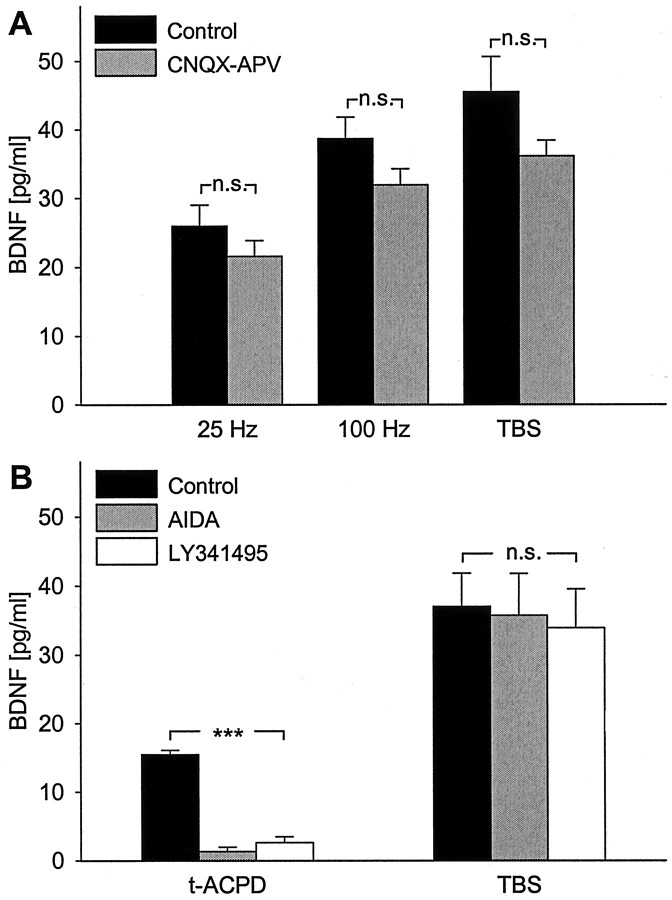
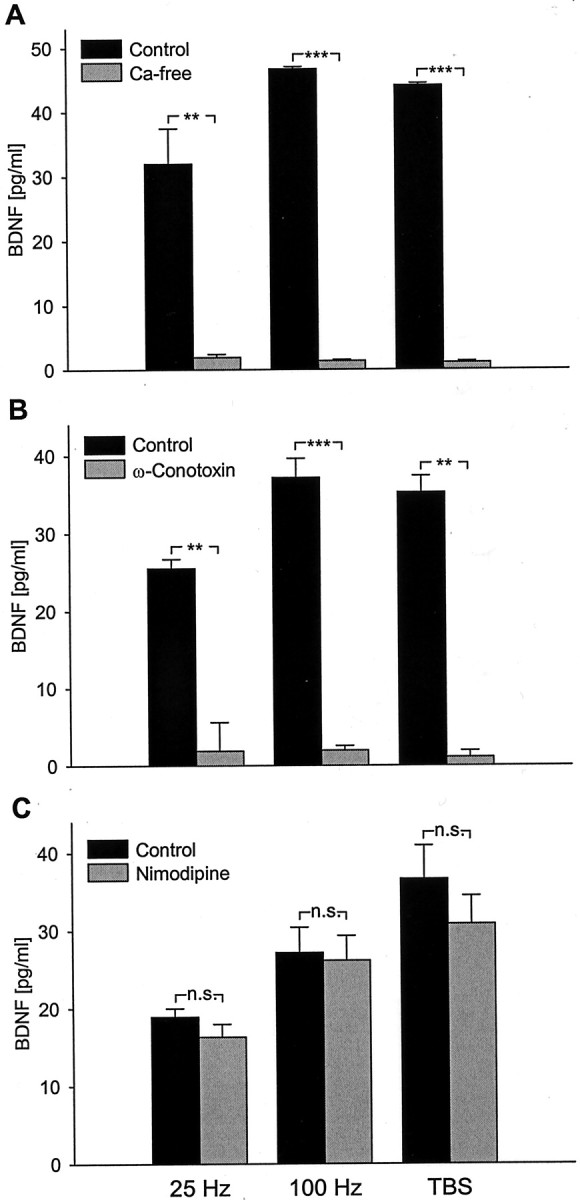
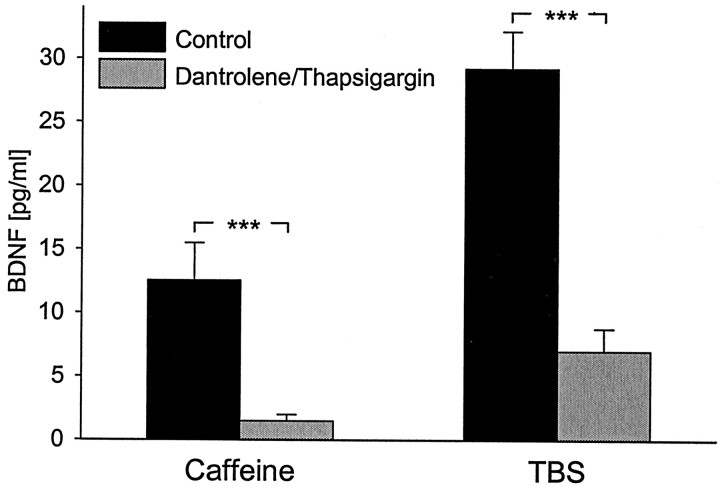
Brain-derived neurotrophic factor (BDNF) plays a critical role in activity-dependent modifications of neuronal connectivity and synaptic strength, including establishment of hippocampal long-term potentiation (LTP). To shed light on mechanisms underlying BDNF-dependent synaptic plasticity, the present study was undertaken to characterize release of native BDNF from newborn rat hippocampal neurons in response to physiologically relevant patterns of electrical field stimulation in culture, including tonic stimulation at 5 Hz, bursting stimulation at 25 and 100 Hz, and theta-burst stimulation (TBS). Release was measured using the ELISA in situ technique, developed in our laboratory to quantify secretion of native BDNF without the need to first overexpress the protein to nonphysiological levels. Each stimulation protocol resulted in a significant increase in BDNF release that was tetrodotoxin sensitive and occurred in the absence of glutamate receptor activation. However, 100 Hz tetanus and TBS, stimulus patterns that are most effective in inducing hippocampal LTP, were significantly more effective in releasing native BDNF than lower-frequency stimulation. For all stimulation protocols tested, removal of extracellular calcium, or blockade of N-type calcium channels, prevented BDNF release. Similarly, depletion of intracellular calcium stores with thapsigargin and treatment with dantrolene, an inhibitor of calcium release from caffeine-ryanodine-sensitive stores, markedly inhibited activity-dependent BDNF release. Our results indicate that BDNF release can encode temporal features of hippocampal neuronal activity. The dual requirement for calcium influx through N-type calcium channels and calcium mobilization from intracellular stores strongly implicates a role for calcium-induced calcium release in activity-dependent BDNF secretion.
Figures
References
-
- Altar CA, Cai N, Bliven T, Juhasz M, Conner JM, Acheson AL, Lindsay RM, Wiegand SJ. Anterograde transport of brain-derived neurotrophic factor and its role in the brain. Nature. 1997;389:856–860. - PubMed
-
- Androutsellis-Theotokis A, McCormack WJ, Bradford HF, Stern GM, Pliego-Rivero FB. The depolarisation-induced release of [125I]BDNF from brain tissue. Brain Res. 1996;743:40–48. - PubMed
-
- Balkowiec A, Katz DM. Cellular mechanisms underlying activity-dependent release of endogenous BDNF from hippocampal neurons. Soc Neurosci Abstr. 2001;27:801.14.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources