

The autosomal recessive hypercholesterolemia (ARH) protein interfaces directly with the clathrin-coat machinery
- PMID: 12451172
- PMCID: PMC138571
- DOI: 10.1073/pnas.252630799
The autosomal recessive hypercholesterolemia (ARH) protein interfaces directly with the clathrin-coat machinery
Abstract
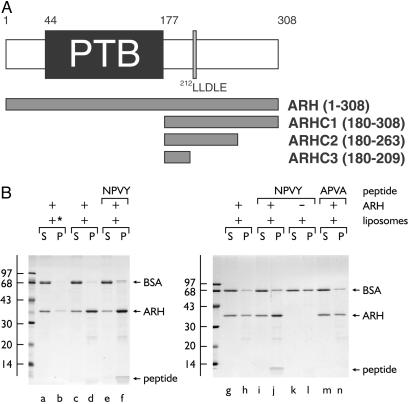
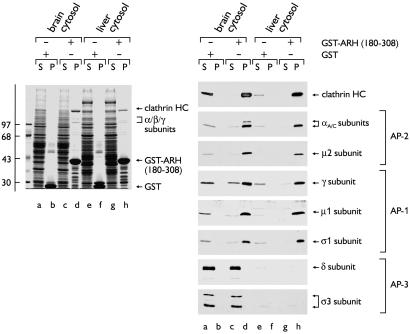
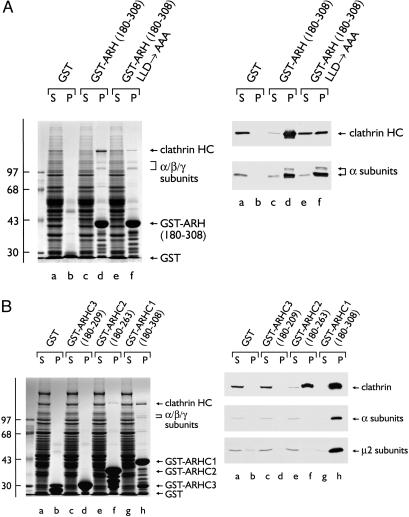
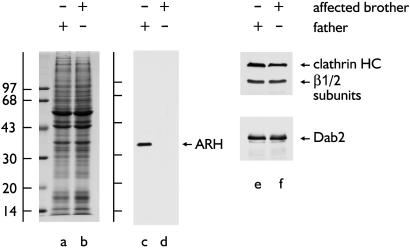
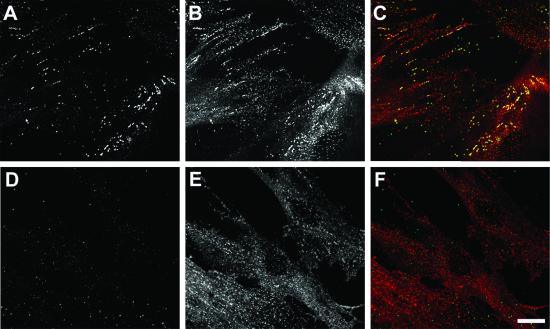
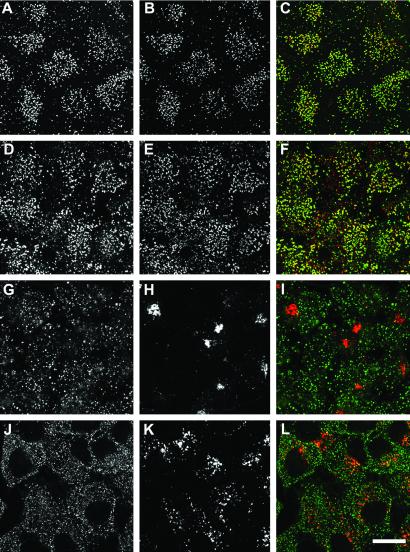
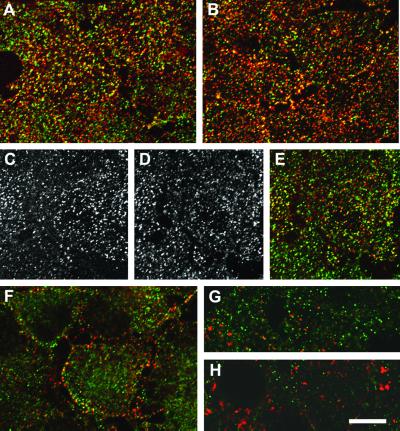
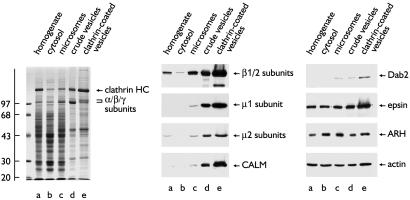
The low density lipoprotein (LDL) receptor plays a pivotal role in cholesterol metabolism. Inherited mutations that disturb the activity of the receptor lead to elevations in plasma cholesterol levels and early-onset coronary atherosclerosis. Defects in either the LDL receptor or apolipoprotein B, the proteinaceous component of LDL particles that binds the LDL receptor, elevate circulating LDL-cholesterol levels in an autosomal-dominant fashion, with heterozygotes displaying values between homozygous and normal individuals. Rarely, similar clinical phenotypes occur with a recessive pattern of inheritance, and several genetic lesions in the autosomal recessive hypercholesterolemia (ARH) gene on chromosome 1 have been mapped in this class of patients. ARH has an N-terminal phosphotyrosine-binding (PTB) domain evolutionarily related to that found in Disabled-2 and numb, two endocytic proteins. PTB domains bind to the consensus sequence FXNPXY, corresponding to the internalization motif of the LDL receptor. We show here that in addition to the FXNPXY sequence, ARH binds directly to soluble clathrin trimers and to clathrin adaptors by a mode involving the independently folded appendage domain of the beta subunit. At steady state, ARH colocalizes with endocytic proteins in HeLa cells, and the LDL receptor fluxes through peripheral ARH-positive sites before delivery to early endosomes. Because ARH also binds directly to phosphoinositides, which regulate clathrin bud assembly at the cell surface, our data suggest that in ARH patients, defective sorting adaptor function in hepatocytes leads to faulty LDL receptor traffic and hypercholesterolemia.
Figures
Similar articles
-
Functional dissection of an AP-2 beta2 appendage-binding sequence within the autosomal recessive hypercholesterolemia protein.J Biol Chem. 2005 May 13;280(19):19270-80. doi: 10.1074/jbc.M501029200. Epub 2005 Feb 22. J Biol Chem. 2005. PMID: 15728179
-
ARH is a modular adaptor protein that interacts with the LDL receptor, clathrin, and AP-2.J Biol Chem. 2002 Nov 15;277(46):44044-9. doi: 10.1074/jbc.M208539200. Epub 2002 Sep 8. J Biol Chem. 2002. PMID: 12221107
-
The modular adaptor protein autosomal recessive hypercholesterolemia (ARH) promotes low density lipoprotein receptor clustering into clathrin-coated pits.J Biol Chem. 2005 Dec 9;280(49):40996-1004. doi: 10.1074/jbc.M509394200. Epub 2005 Sep 22. J Biol Chem. 2005. PMID: 16179341
-
Genetics, clinical phenotype, and molecular cell biology of autosomal recessive hypercholesterolemia.Arterioscler Thromb Vasc Biol. 2003 Nov 1;23(11):1963-70. doi: 10.1161/01.ATV.0000094410.66558.9A. Epub 2003 Sep 4. Arterioscler Thromb Vasc Biol. 2003. PMID: 12958046 Review.
-
Autosomal recessive hypercholesterolemia.Semin Vasc Med. 2004 Aug;4(3):241-8. doi: 10.1055/s-2004-861491. Semin Vasc Med. 2004. PMID: 15630633 Review.
Cited by
-
Analysis of the AP-2 adaptor complex and cargo during clathrin-mediated endocytosis.Traffic. 2005 Jul;6(7):539-47. doi: 10.1111/j.1600-0854.2005.00280.x. Traffic. 2005. PMID: 15941406 Free PMC article.
-
Regulation of ApoB secretion by the low density lipoprotein receptor requires exit from the endoplasmic reticulum and interaction with ApoE or ApoB.J Biol Chem. 2008 Apr 25;283(17):11374-81. doi: 10.1074/jbc.M710457200. Epub 2008 Feb 13. J Biol Chem. 2008. PMID: 18272520 Free PMC article.
-
Function and dysfunction of the PI system in membrane trafficking.EMBO J. 2008 Oct 8;27(19):2457-70. doi: 10.1038/emboj.2008.169. Epub 2008 Sep 11. EMBO J. 2008. PMID: 18784754 Free PMC article. Review.
-
Weak Molecular Interactions in Clathrin-Mediated Endocytosis.Front Mol Biosci. 2017 Nov 14;4:72. doi: 10.3389/fmolb.2017.00072. eCollection 2017. Front Mol Biosci. 2017. PMID: 29184887 Free PMC article. Review.
-
S-Nitrosylation: Mechanistic Links between Nitric Oxide Signaling and Atherosclerosis.Curr Atheroscler Rep. 2025 Aug 7;27(1):78. doi: 10.1007/s11883-025-01321-y. Curr Atheroscler Rep. 2025. PMID: 40772994 Review.
References
-
- Chen W. J., Goldstein, J. L. & Brown, M. S. (1990) J. Biol. Chem. 265, 3116-3123. - PubMed
-
- Davis C. G., Lehrman, M. A., Russell, D. W., Anderson, R. G., Brown, M. S. & Goldstein, J. L. (1986) Cell 45, 15-24. - PubMed
-
- Warren R. A., Green, F. A., Stenberg, P. E. & Enns, C. A. (1998) J. Biol. Chem. 273, 17056-17063. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
