

Evidence for lateral transfer of genes encoding ferredoxins, nitroreductases, NADH oxidase, and alcohol dehydrogenase 3 from anaerobic prokaryotes to Giardia lamblia and Entamoeba histolytica
- PMID: 12455953
- PMCID: PMC118039
- DOI: 10.1128/EC.1.2.181-190.2002
Evidence for lateral transfer of genes encoding ferredoxins, nitroreductases, NADH oxidase, and alcohol dehydrogenase 3 from anaerobic prokaryotes to Giardia lamblia and Entamoeba histolytica
Abstract
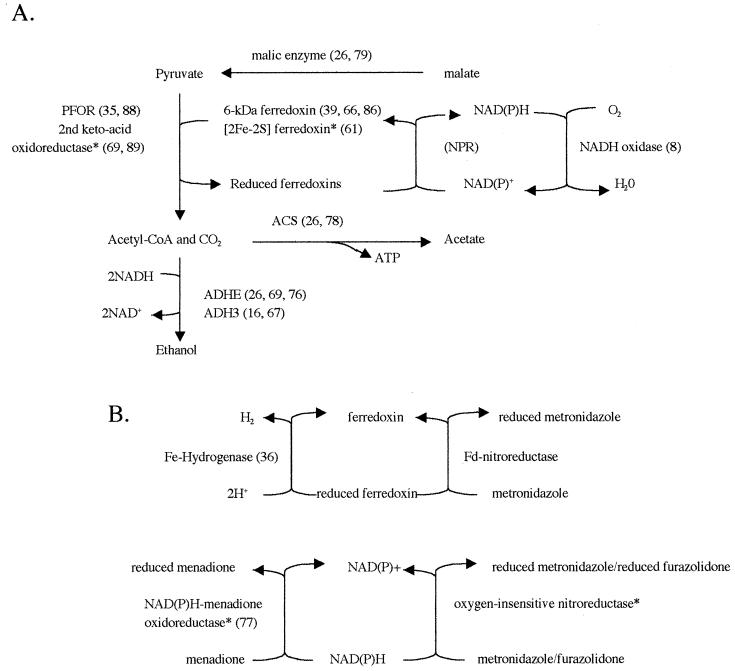
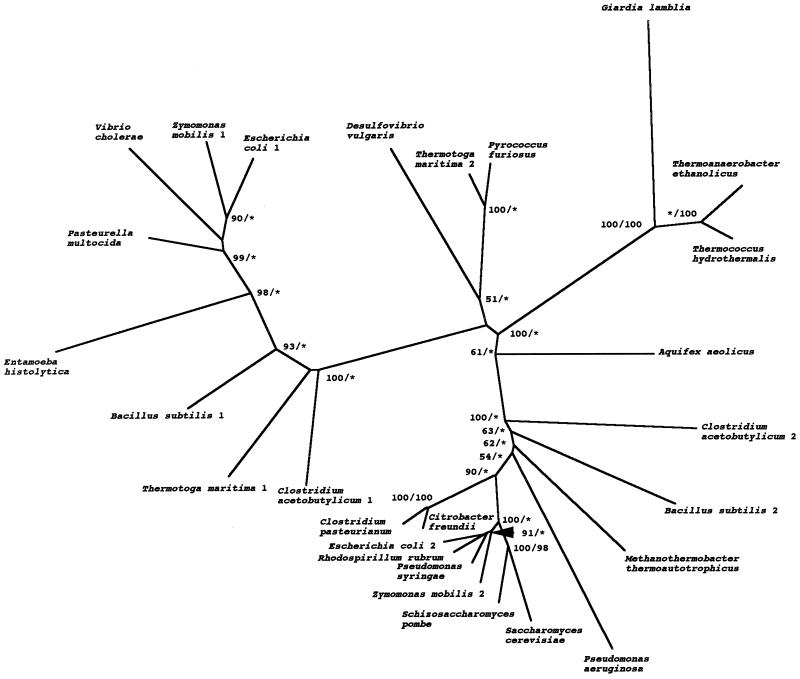
Giardia lamblia and Entamoeba histolytica are amitochondriate, microaerophilic protists which use fermentation enzymes like those of bacteria to survive anaerobic conditions within the intestinal lumen. Genes encoding fermentation enzymes and related electron transport peptides (e.g., ferredoxins) in giardia organisms and amebae are hypothesized to be derived from either an ancient anaerobic eukaryote (amitochondriate fossil hypothesis), a mitochondrial endosymbiont (hydrogen hypothesis), or anaerobic bacteria (lateral transfer hypothesis). The goals here were to complete the molecular characterization of giardial and amebic fermentation enzymes and to determine the origins of the genes encoding them, when possible. A putative giardia [2Fe-2S]ferredoxin which had a hypothetical organelle-targeting sequence at its N terminus showed similarity to mitochondrial ferredoxins and the hydrogenosomal ferredoxin of Trichomonas vaginalis (another luminal protist). However, phylogenetic trees were star shaped, with weak bootstrap support, so we were unable to confirm or rule out the endosymbiotic origin of the giardia [2Fe-2S]ferredoxin gene. Putative giardial and amebic 6-kDa ferredoxins, ferredoxin-nitroreductase fusion proteins, and oxygen-insensitive nitroreductases each tentatively supported the lateral transfer hypothesis. Although there were not enough sequences to perform meaningful phylogenetic analyses, the unique common occurrence of these peptides and enzymes in giardia organisms, amebae, and the few anaerobic prokaryotes suggests the possibility of lateral transfer. In contrast, there was more robust phylogenetic evidence for the lateral transfer of G. lamblia genes encoding an NADH oxidase from a gram-positive coccus and a microbial group 3 alcohol dehydrogenase from thermoanaerobic prokaryotes. In further support of lateral transfer, the G. lamblia NADH oxidase and adh3 genes appeared to have an evolutionary history distinct from those of E. histolytica.
Figures



References
-
- Adams, M. D., S. E. Celniker, R. A. Holt, et al. 2000.. The genome sequence of Drosophila melanogaster. Science 287:2185-2195. - PubMed
-
- Andersson, S. G. E., and C. G. Kurland. 1999. Origins of mitochondria and hydrogenosomes. Curr. Opin. Microbiol. 2:535-541. - PubMed
-
- Anonymous. 1998. Genome sequence of the nematode C. elegans: a platform for investigating biology. Science 282:2012-2018. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
