

Nnf1p, Dsn1p, Mtw1p, and Nsl1p: a new group of proteins important for chromosome segregation in Saccharomyces cerevisiae
- PMID: 12455957
- PMCID: PMC118027
- DOI: 10.1128/EC.1.2.229-240.2002
Nnf1p, Dsn1p, Mtw1p, and Nsl1p: a new group of proteins important for chromosome segregation in Saccharomyces cerevisiae
Abstract
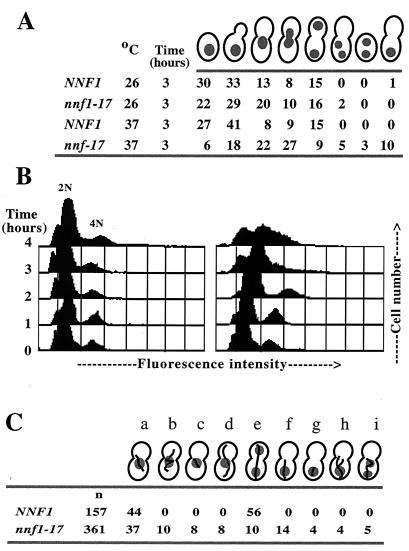
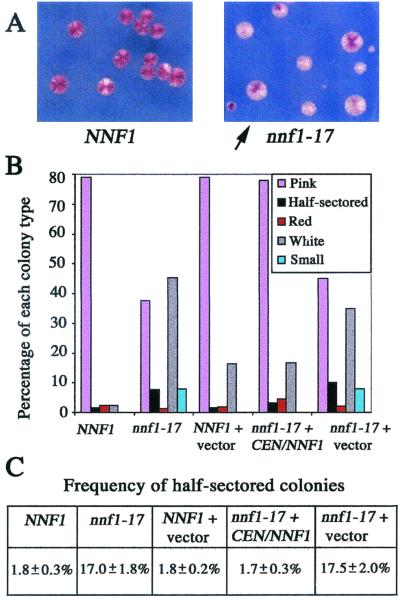
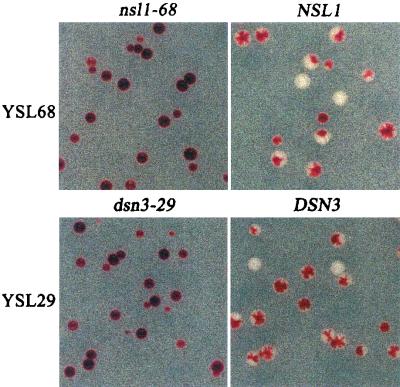
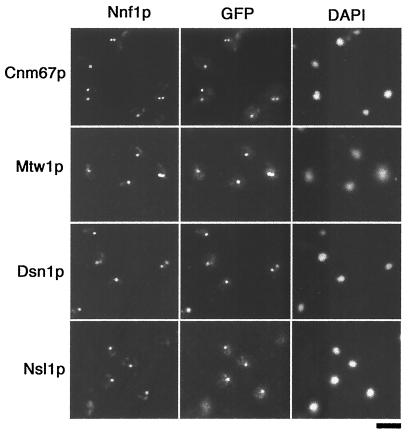
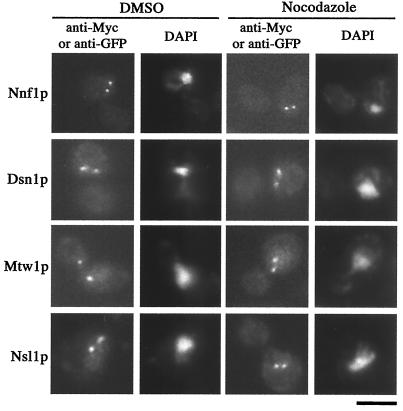
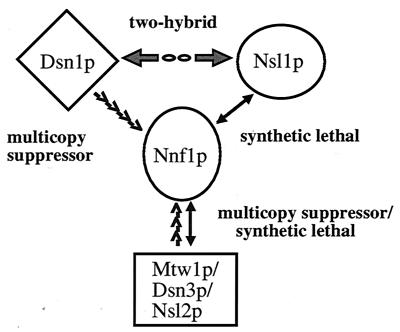
Previously, antibodies were raised against a nuclear envelope-enriched fraction of yeast, and the essential gene NNF1 was cloned by reverse genetics. Here it is shown that the conditional nnf1-17 mutant has decreased stability of a minichromosome in addition to mitotic spindle defects. I have identified the novel essential genes DSN1, DSN3, and NSL1 through genetic interactions with nnf1-17. Dsn3p was found to be equivalent to the kinetochore protein Mtw1p. By indirect immunofluorescence, all four proteins, Nnf1p, Mtw1p, Dsn1p, and Nsl1p, colocalize and are found in the region of the spindle poles. Based on the colocalization of these four proteins, the minichromosome instability and the spindle defects seen in nnf1 mutants, I propose that Nnf1p is part of a new group of proteins necessary for chromosome segregation.
Figures

Similar articles
-
Nsl1p is essential for the establishment of bipolarity and the localization of the Dam-Duo complex.EMBO J. 2003 Dec 15;22(24):6584-97. doi: 10.1093/emboj/cdg636. EMBO J. 2003. PMID: 14657030 Free PMC article.
-
Interactions between centromere complexes in Saccharomyces cerevisiae.Mol Biol Cell. 2003 Dec;14(12):4931-46. doi: 10.1091/mbc.e03-06-0419. Epub 2003 Oct 17. Mol Biol Cell. 2003. PMID: 14565975 Free PMC article.
-
An Mtw1 complex promotes kinetochore biorientation that is monitored by the Ipl1/Aurora protein kinase.Dev Cell. 2003 Nov;5(5):735-45. doi: 10.1016/s1534-5807(03)00322-8. Dev Cell. 2003. PMID: 14602074
-
Protein kinases in mitotic phosphorylation of budding yeast CENP-A.Curr Genet. 2019 Dec;65(6):1325-1332. doi: 10.1007/s00294-019-00997-5. Epub 2019 May 22. Curr Genet. 2019. PMID: 31119371 Review.
-
A one-sided view of kinetochore attachment in meiosis.Cell. 2006 Sep 22;126(6):1030-2. doi: 10.1016/j.cell.2006.09.005. Cell. 2006. PMID: 16990129 Review.
Cited by
-
Cdc14-dependent dephosphorylation of a kinetochore protein prior to anaphase in Saccharomyces cerevisiae.Genetics. 2010 Dec;186(4):1487-91. doi: 10.1534/genetics.110.123653. Epub 2010 Oct 5. Genetics. 2010. PMID: 20923974 Free PMC article.
-
A system for enhancing genome-wide coexpression dynamics study.Proc Natl Acad Sci U S A. 2004 Nov 2;101(44):15561-6. doi: 10.1073/pnas.0402962101. Epub 2004 Oct 18. Proc Natl Acad Sci U S A. 2004. PMID: 15492223 Free PMC article.
-
Methylation of CENP-A/Cse4 on arginine 143 and lysine 131 regulates kinetochore stability in yeast.Genetics. 2023 Apr 6;223(4):iyad028. doi: 10.1093/genetics/iyad028. Genetics. 2023. PMID: 36810679 Free PMC article.
-
The conserved Spc7 protein is required for spindle integrity and links kinetochore complexes in fission yeast.Mol Biol Cell. 2007 Jul;18(7):2441-54. doi: 10.1091/mbc.e06-08-0738. Epub 2007 Apr 18. Mol Biol Cell. 2007. PMID: 17442892 Free PMC article.
-
Leveraging machine learning essentiality predictions and chemogenomic interactions to identify antifungal targets.Nat Commun. 2021 Nov 11;12(1):6497. doi: 10.1038/s41467-021-26850-3. Nat Commun. 2021. PMID: 34764269 Free PMC article.
References
-
- Boeke, J. D., J. Trueheart, G. Natsoulis, and G. R. Fink. 1987. 5-Fluoroorotic acid as a selective agent in yeast molecular genetics. Methods Enzymol. 154:164-175. - PubMed
-
- Byers, B., and L. Goetsch. 1974. Duplication of spindle plaques and integration of the yeast cell cycle. Cold Spring Harbor Symp. Quant. Biol. 38:123-131. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
