

Sgt1p contributes to cyclic AMP pathway activity and physically interacts with the adenylyl cyclase Cyr1p/Cdc35p in budding yeast
- PMID: 12456005
- PMCID: PMC118006
- DOI: 10.1128/EC.1.4.568-582.2002
Sgt1p contributes to cyclic AMP pathway activity and physically interacts with the adenylyl cyclase Cyr1p/Cdc35p in budding yeast
Abstract
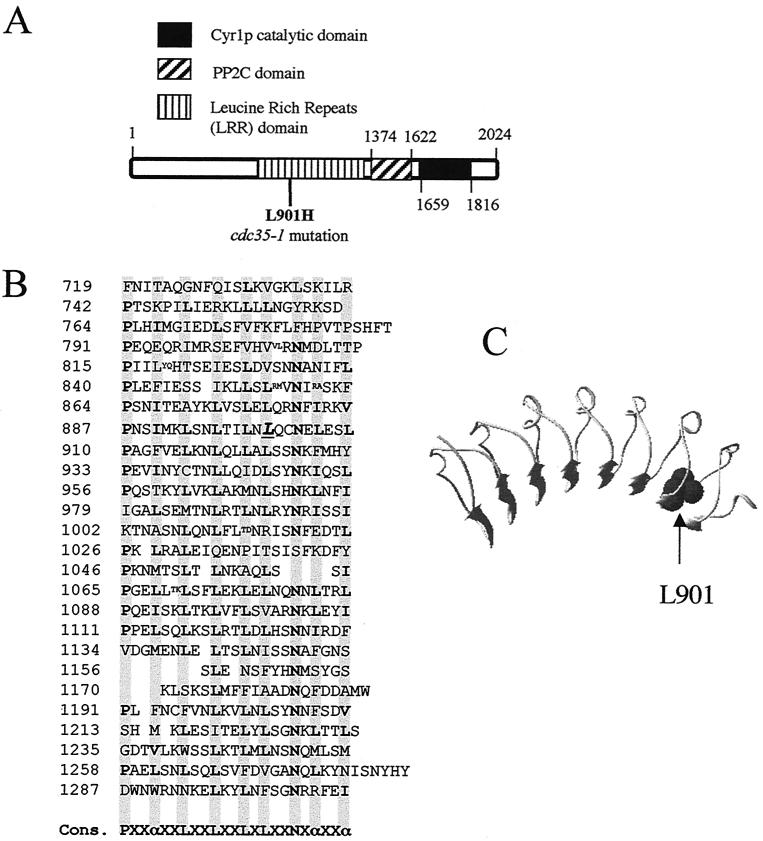
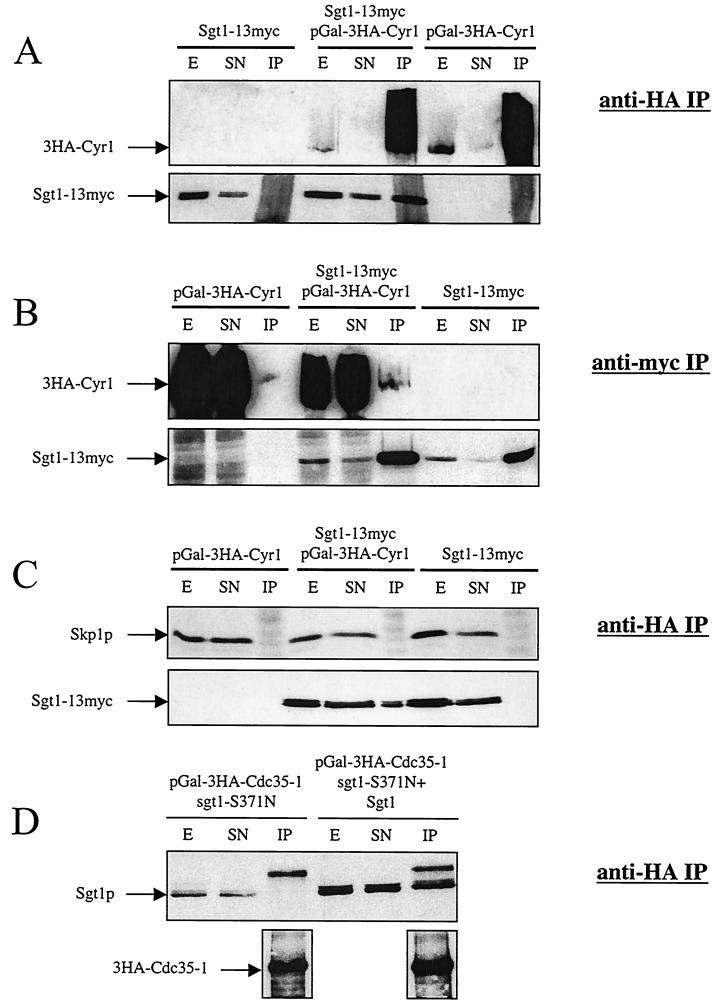
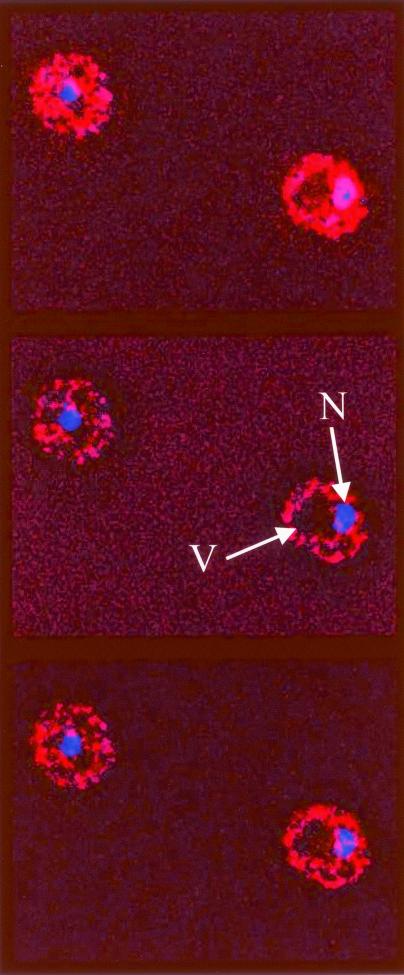
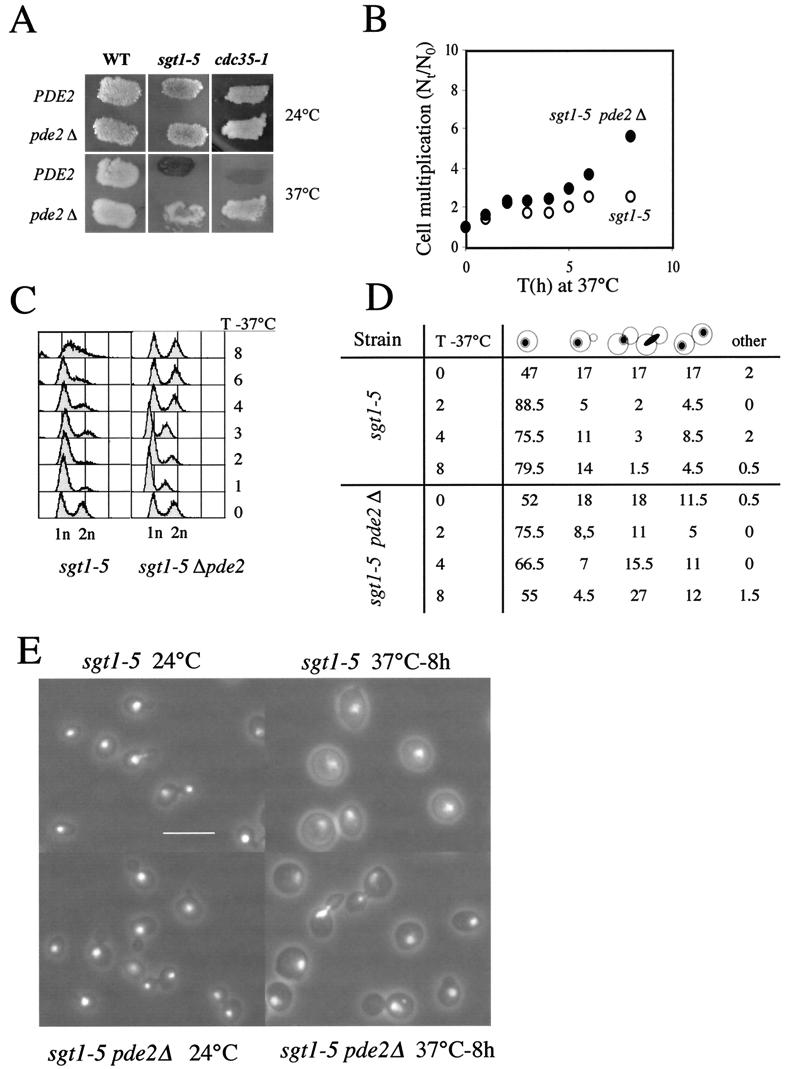
Sgt1p is a highly conserved eucaryotic protein that is required for both SCF (Skp1p/Cdc53p-Cullin-F-box)-mediated ubiquitination and kinetochore function in yeast. We show here that Sgtlp is also involved in the cyclic AMP (cAMP) pathway in Saccharomyces cerevisiae. SGT1 is an allele-specific suppressor of cdc35-1, a thermosensitive mutation in the leucine-rich repeat domain of the adenylyl cyclase Cyrlp/Cdc35p. We demonstrate that Sgt1p and Cyrlp/Cdc35p physically interact and that the activity of the cAMP pathway is affected in an sgt1 conditional mutant. Sequence analysis suggests that Sgtlp has features of a cochaperone. Thus, Sgt1p is a novel activator of adenylyl cyclase in S. cerevisiae and may function in the assembly or the conformational activation of specific multiprotein complexes.
Figures



Similar articles
-
Schizosaccharomyces pombe Git7p, a member of the Saccharomyces cerevisiae Sgtlp family, is required for glucose and cyclic AMP signaling, cell wall integrity, and septation.Eukaryot Cell. 2002 Aug;1(4):558-67. doi: 10.1128/EC.1.4.558-567.2002. Eukaryot Cell. 2002. PMID: 12456004 Free PMC article.
-
Sgt1p is a unique co-chaperone that acts as a client adaptor to link Hsp90 to Skp1p.J Biol Chem. 2006 Nov 3;281(44):33739-48. doi: 10.1074/jbc.M603847200. Epub 2006 Aug 31. J Biol Chem. 2006. PMID: 16945921
-
The interaction between Sgt1p and Skp1p is regulated by HSP90 chaperones and is required for proper CBF3 assembly.Mol Cell Biol. 2004 Oct;24(20):8938-50. doi: 10.1128/MCB.24.20.8938-8950.2004. Mol Cell Biol. 2004. PMID: 15456868 Free PMC article.
-
Novel sensing mechanisms and targets for the cAMP-protein kinase A pathway in the yeast Saccharomyces cerevisiae.Mol Microbiol. 1999 Sep;33(5):904-18. doi: 10.1046/j.1365-2958.1999.01538.x. Mol Microbiol. 1999. PMID: 10476026 Review.
-
Phosphate homeostasis in the yeast Saccharomyces cerevisiae, the key role of the SPX domain-containing proteins.FEBS Lett. 2012 Feb 17;586(4):289-95. doi: 10.1016/j.febslet.2012.01.036. Epub 2012 Jan 27. FEBS Lett. 2012. PMID: 22285489 Review.
Cited by
-
Glucose Signaling Is Connected to Chromosome Segregation Through Protein Kinase A Phosphorylation of the Dam1 Kinetochore Subunit in Saccharomycescerevisiae.Genetics. 2019 Feb;211(2):531-547. doi: 10.1534/genetics.118.301727. Epub 2018 Dec 13. Genetics. 2019. PMID: 30546002 Free PMC article.
-
Identification of an Hsp90 mutation that selectively disrupts cAMP/PKA signaling in Saccharomyces cerevisiae.Curr Genet. 2012 Jun;58(3):149-63. doi: 10.1007/s00294-012-0373-7. Epub 2012 Mar 30. Curr Genet. 2012. PMID: 22461145
-
SUGT1 is a prognostic biomarker and is associated with immune infiltrates in ovarian cancer.Eur J Med Res. 2025 Jan 10;30(1):21. doi: 10.1186/s40001-024-02232-5. Eur J Med Res. 2025. PMID: 39794869 Free PMC article.
-
Comprehensive Interactome Analysis for the Sole Adenylyl Cyclase Cyr1 of Candida albicans.Microbiol Spectr. 2022 Dec 21;10(6):e0393422. doi: 10.1128/spectrum.03934-22. Epub 2022 Oct 31. Microbiol Spectr. 2022. PMID: 36314909 Free PMC article.
-
Application of cDNA array for studying the gene expression profile of mature appressoria of Magnaporthe grisea.J Zhejiang Univ Sci B. 2007 Feb;8(2):88-97. doi: 10.1631/jzus.2007.B0088. J Zhejiang Univ Sci B. 2007. PMID: 17266183 Free PMC article.
References
-
- Bai, C., P. Sen, K. Hofmann, L. Ma, M. Goebl, J. W. Harper, and S. J. Elledge. 1996. SKP1 connects cell cycle regulators to the ubiquitin proteolysis machinery through a novel motif, the F-box. Cell 86:263-274. - PubMed
-
- Baroni, M. D., P. Monti, and L. Alberghina. 1994. Repression of growth-regulated G1 cyclin expression by cyclic AMP in budding yeast. Nature 371:339-342. - PubMed
-
- Beckner, S. K., S. Hattori, and T. Y. Shih. 1985. The ras oncogene product p21 is not a regulatory component of adenylate cyclase. Nature 317:71-72. - PubMed
-
- Beebe, S. J. 1994. The cAMP-dependent protein kinases and cAMP signal transduction. Semin. Cancer Biol. 5:285-294. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
