

The contribution of intracellular calcium stores to mEPSCs recorded in layer II neurones of rat barrel cortex
- PMID: 12456831
- PMCID: PMC2290677
- DOI: 10.1113/jphysiol.2002.022103
The contribution of intracellular calcium stores to mEPSCs recorded in layer II neurones of rat barrel cortex
Abstract
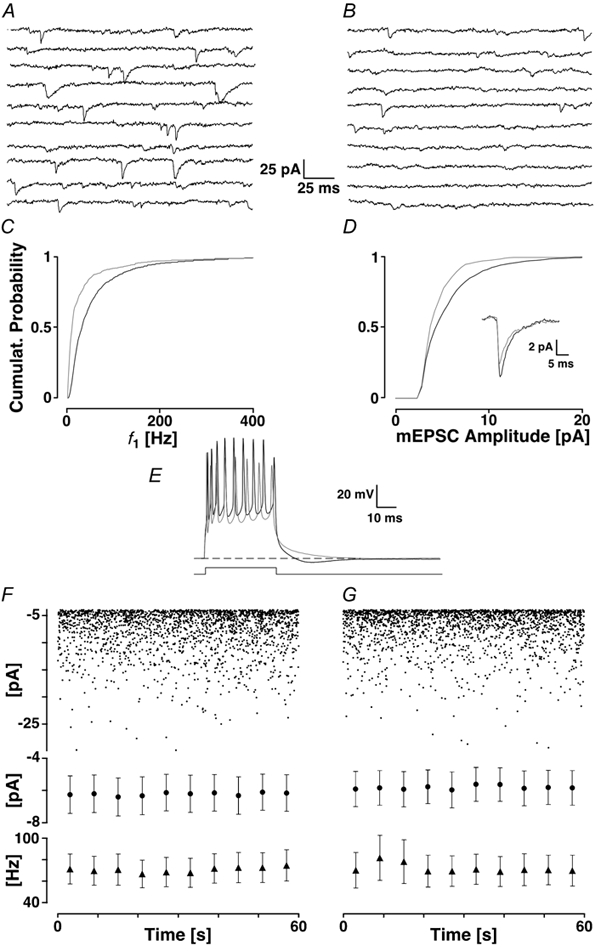
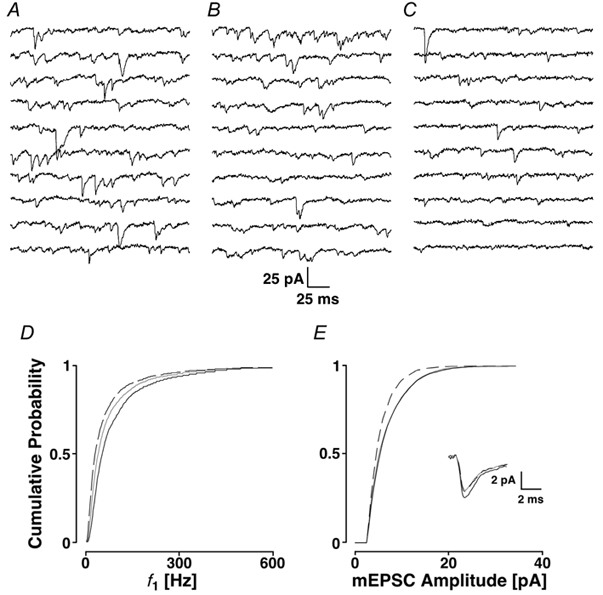
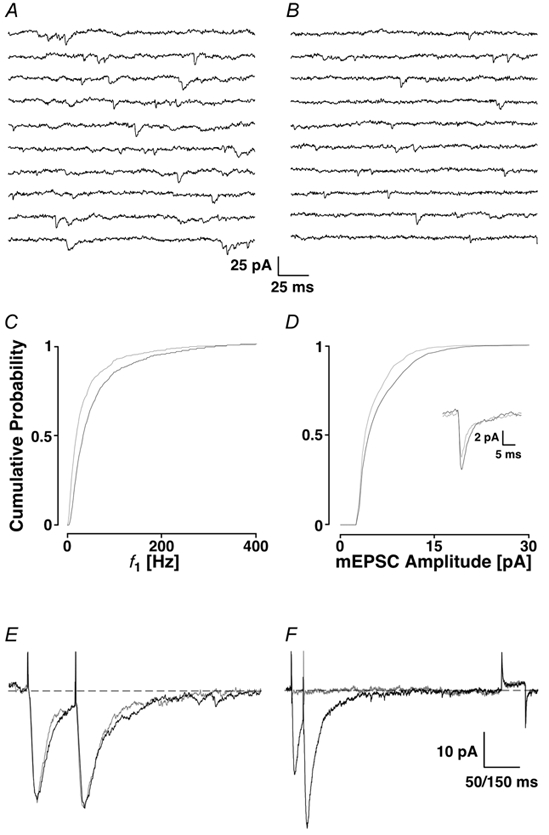
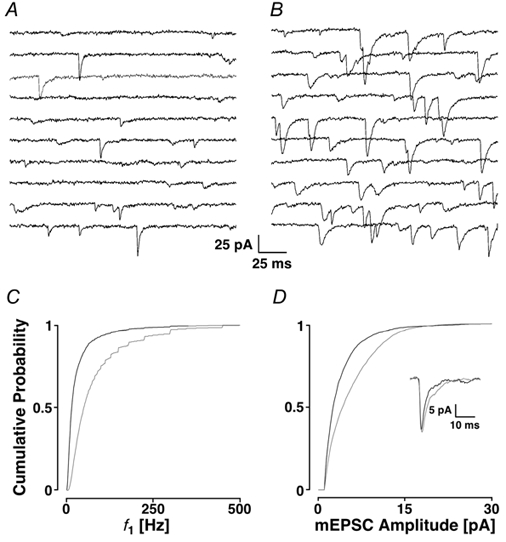
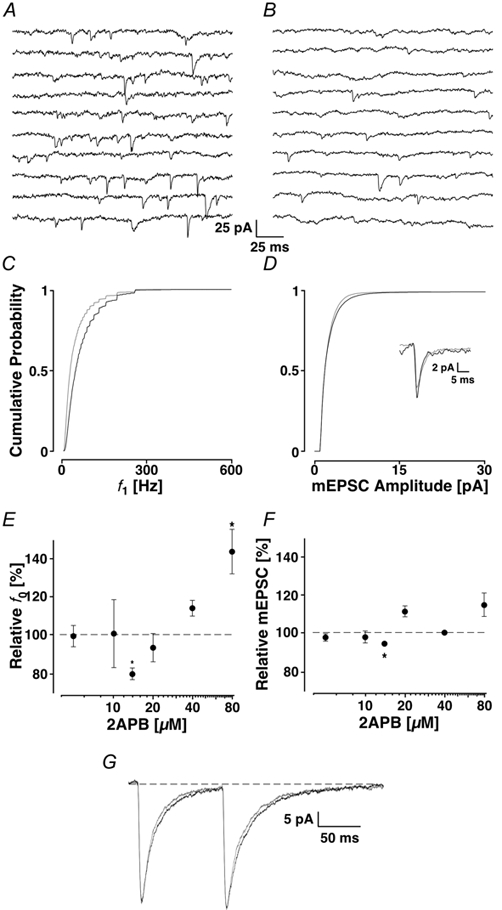
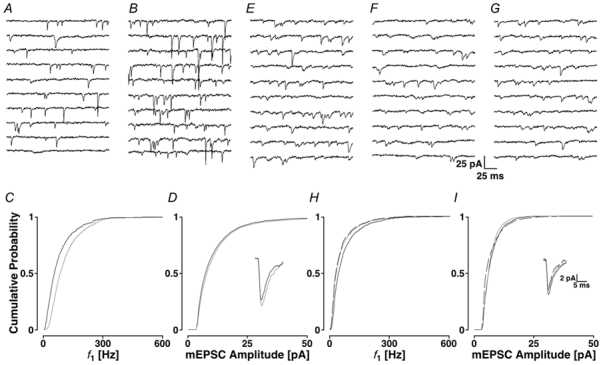
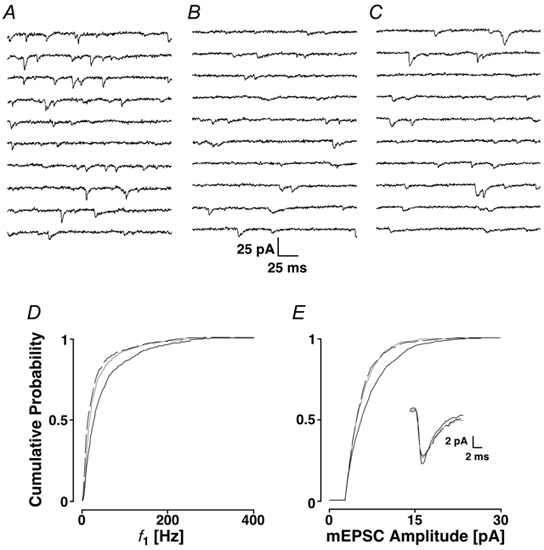
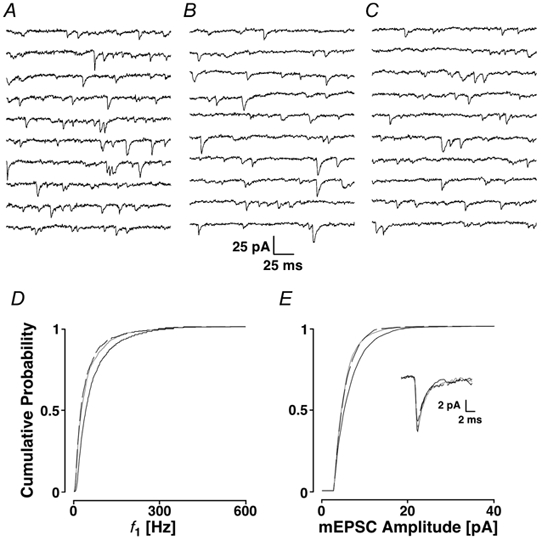
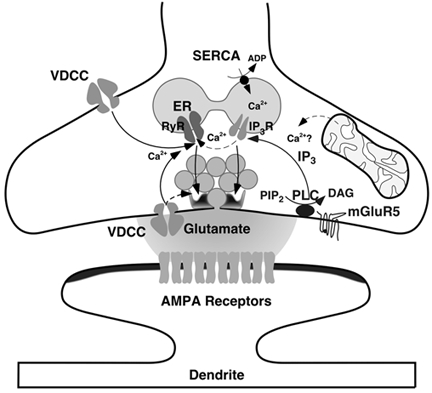
Loading slices of rat barrel cortex with 50 microM BAPTA-AM while recording from pyramidal cells in layer II induces a marked reduction in both the frequency and amplitudes of mEPSCs. These changes are due to a presynaptic action. Blocking the refilling of Ca(2+) stores with 20 microM cyclopiazonic acid (CPA), a SERCA pump inhibitor, in conjunction with neuronal depolarisation to activate Ca(2+) stores, results in a similar reduction of mEPSCs to that observed with BAPTA-AM, indicating that the source for intracellular Ca(2+) is the endoplasmic reticulum. Block or activation of ryanodine receptors by 20 microM ryanodine or 10 mM caffeine, respectively, shows that a significant proportion of mEPSCs are caused by Ca(2+) release from ryanodine stores. Blocking IP(3) receptors with 14 microM 2-aminoethoxydiphenylborane (2APB) also reduces the frequency and amplitude of mEPSCs, indicating the involvement of IP(3) stores in the generation of mEPSCs. Activation of group I metabotropic receptors with 20 microM (RS)-3,5-dihydroxyphenylglycine (DHPG) results in a significant increase in the frequency of mEPSCs, further supporting the role of IP(3) receptors and indicating a role of group I metabotropic receptors in causing transmitter release. Statistical evidence is presented for Ca(2+)-induced Ca(2+) release (CICR) from ryanodine stores after the spontaneous opening of IP(3) stores.
Figures
References
-
- Bird MM. Presynaptic and postsynaptic organelles of synapses formed in cultures of previously dissociated mouse spinal cord. Cell and Tissue Research. 1978;194:503–511. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous