

An mRNA structure that controls gene expression by binding FMN
- PMID: 12456892
- PMCID: PMC138538
- DOI: 10.1073/pnas.212628899
An mRNA structure that controls gene expression by binding FMN
Abstract
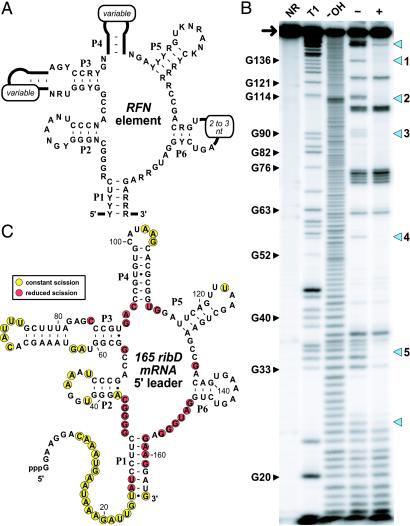
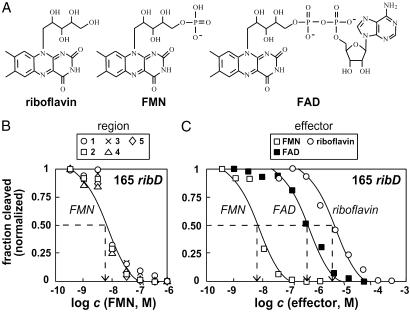
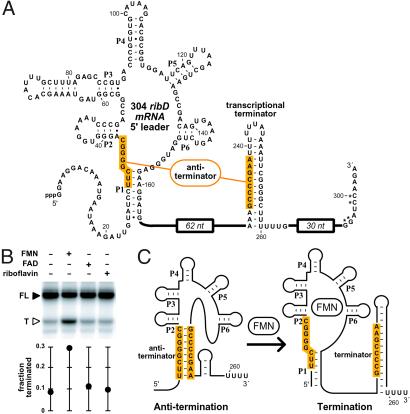
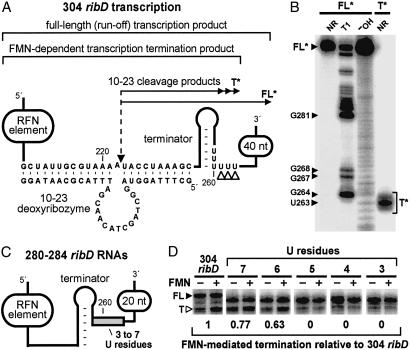
The RFN element is a highly conserved domain that is found frequently in the 5'-untranslated regions of prokaryotic mRNAs that encode for flavin mononucleotide (FMN) biosynthesis and transport proteins. We report that this domain serves as the receptor for a metabolite-dependent riboswitch that directly binds FMN in the absence of proteins. Our results also indicate that in Bacillus subtilis, the riboswitch most likely controls gene expression by causing premature transcription termination of the ribDEAHT operon and precluding access to the ribosome-binding site of ypaA mRNA. Sequence and structural analyses indicate that the RFN element is a natural FMN-binding aptamer, the allosteric character of which is harnessed to control gene expression.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
