

CD44 is required for two consecutive steps in HGF/c-Met signaling
- PMID: 12464636
- PMCID: PMC187488
- DOI: 10.1101/gad.242602
CD44 is required for two consecutive steps in HGF/c-Met signaling
Abstract
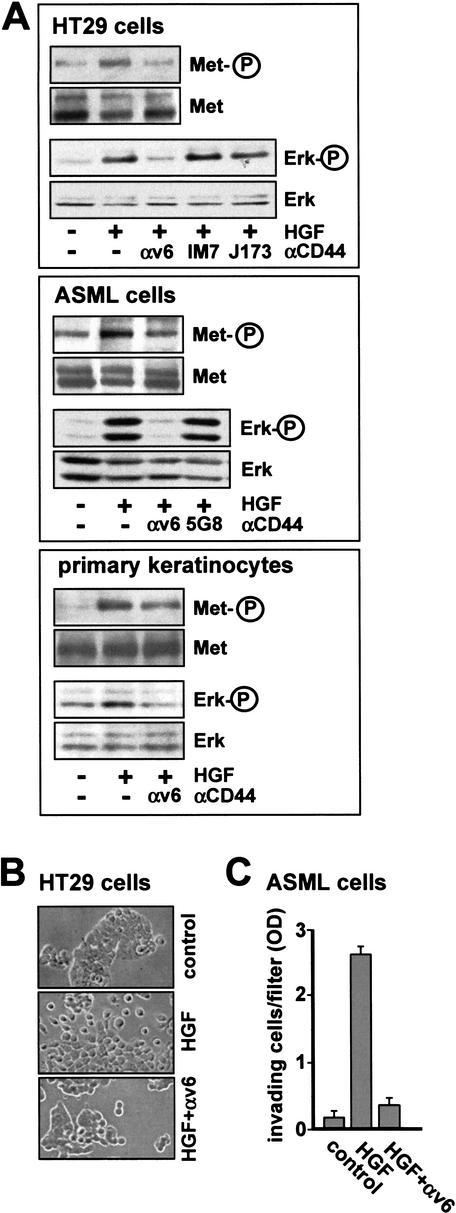
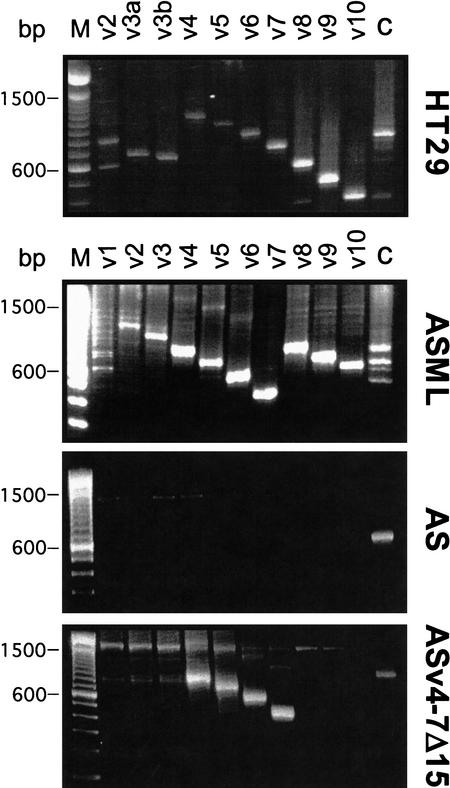
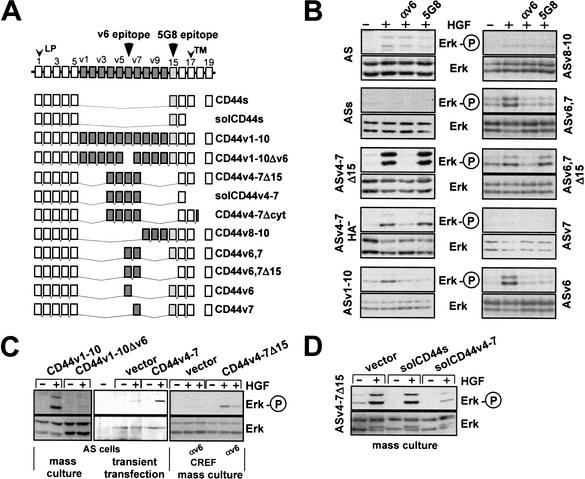
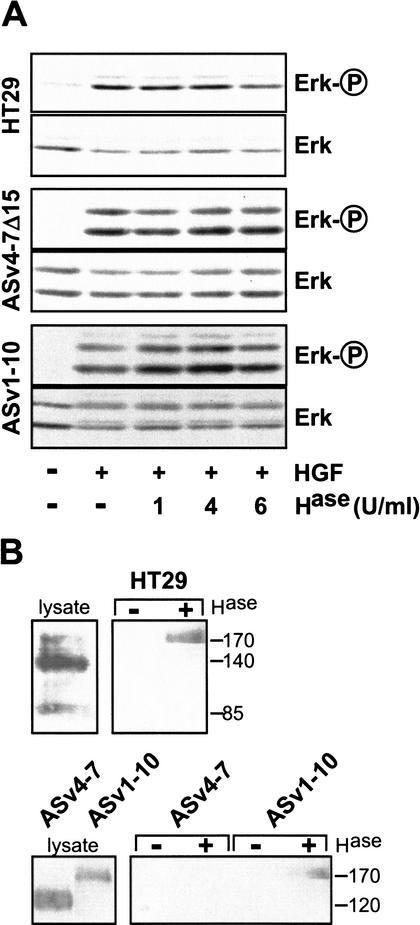
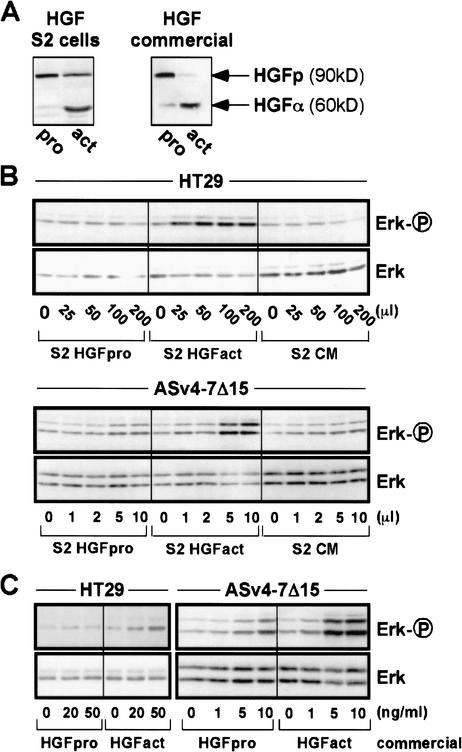
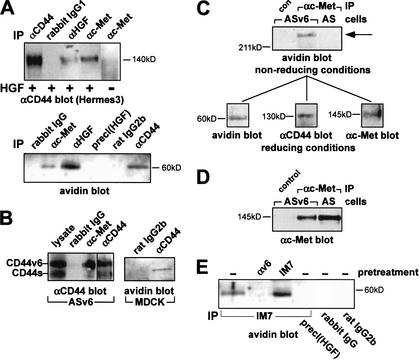
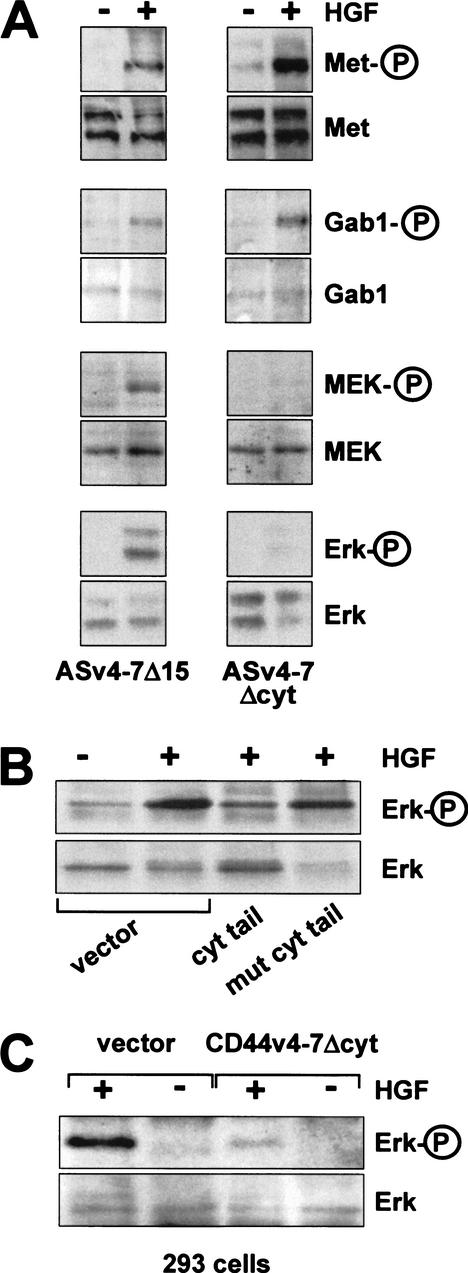
The tyrosine kinase receptor c-Met and its ligand HGF/SF, ezrin, and splice variants of CD44 have independently been identified as tumor metastasis-associated proteins. We now show that these proteins cooperate. A CD44 isoform containing variant exon v6 sequences is strictly required for c-Met activation by HGF/SF in rat and human carcinoma cells, in established cell lines as well as in primary keratinocytes. CD44v6-deficient tumor cells were unable to activate c-Met unless they were transfected with a CD44v6-bearing isoform. Antibodies to two v6-encoded epitopes inhibited autophosphorylation of c-Met by interfering with the formation of a complex formed by c-Met, CD44v6, and HGF/SF. In addition, signal transduction from activated c-Met to MEK and Erk required the presence of the cytoplasmic tail of CD44 including a binding motif for ERM proteins. This suggests a role for ERM proteins and possibly their link to the cortical actin cytoskeleton in signal transfer.
Figures
References
-
- Bardelli A, Comoglio PM. Scatter factor receptors are key players in a unique multistep program leading to invasive growth. Ciba Found Symp. 1997;212:133–144. - PubMed
-
- Bardelli A, Pugliese L, Comoglio PM. “Invasive-growth” signaling by the Met/HGF receptor: The hereditary renal carcinoma connection. Biochim Biophys Acta. 1997;1333:41–51. - PubMed
-
- Birchmeier C, Gherardi E. Developmental roles of HGF/SF and its receptor, the c-Met tyrosine kinase. Trends Cell Biol. 1998;8:404–410. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous