

Evidence for a fast, intrachromosomal conversion mechanism from mapping of nucleotide variants within a homogeneous alpha-satellite DNA array
- PMID: 12466285
- PMCID: PMC187568
- DOI: 10.1101/gr.451502
Evidence for a fast, intrachromosomal conversion mechanism from mapping of nucleotide variants within a homogeneous alpha-satellite DNA array
Abstract
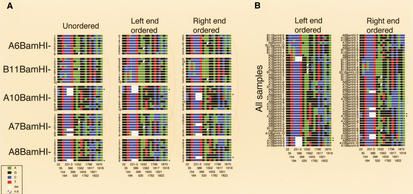
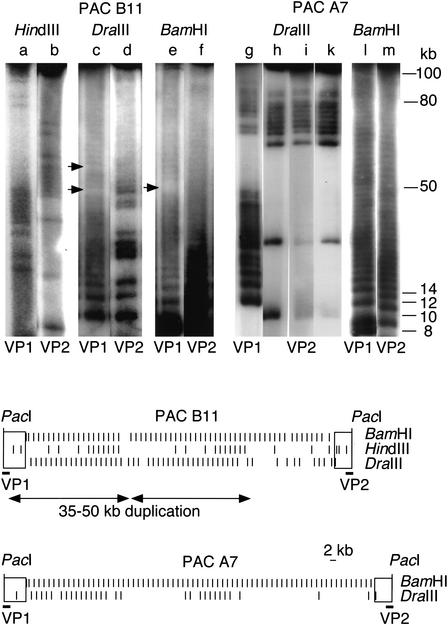
Assuming that patterns of sequence variants within highly homogeneous centromeric tandem repeat arrays can tell us which molecular turnover mechanisms are presently at work, we analyzed the alpha-satellite tandem repeat array DXZ1 of one human X chromosome. Here we present accurate snapshots from this dark matter of the genome. We demonstrate stable and representative cloning of the array in a P1 artificial chromosome (PAC) library, use samples of higher-order repeats subcloned from five unmapped PACs (120-160 kb) to identify common variants, and show that such variants are presently in a fixed transition state. To characterize patterns of variant spread throughout homogeneous array segments, we use a novel partial restriction and pulsed-field gel electrophoresis mapping approach. We find an older large-scale (35-50 kb) duplication event supporting the evolutionarily important unequal crossing-over hypothesis, but generally find independent variant occurrence and a paucity of potential de novo mutations within segments of highest homogeneity (99.1%-99.3%). Within such segments, a highly nonrandom variant clustering within adjacent higher-order repeats was found in the absence of haplotypic repeats. Such variant clusters are hardly explained by interchromosomal, fixation-driving mechanisms and likely reflect a fast, localized, intrachromosomal sequence conversion mechanism.
Figures
References
-
- Alexandrov IA, Mitkevich SP, Yurov YB. The phylogeny of human chromosome specific α satellites. Chromosoma. 1988;96:443–453. - PubMed
-
- Barry AE, Howman EV, Cancilla MR, Saffery R, Choo KHA. Sequence analysis of an 80 kb human neocentromere. Hum Mol Genet. 1999;8:217–227. - PubMed
-
- Bollag RJ, Waldman AS, Liskay RM. Homologous recombination in mammalian cells. Annu Rev Genet. 1989;23:199–225. - PubMed
-
- Charlesworth B, Sniegowski P, Stephan W. The evolutionary dynamics of repetitive DNA in eukaryotes. Nature. 1994;371:215–220. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources