

Plasmodium falciparum erythrocyte invasion through glycophorin C and selection for Gerbich negativity in human populations
- PMID: 12469115
- PMCID: PMC3728825
- DOI: 10.1038/nm807
Plasmodium falciparum erythrocyte invasion through glycophorin C and selection for Gerbich negativity in human populations
Abstract
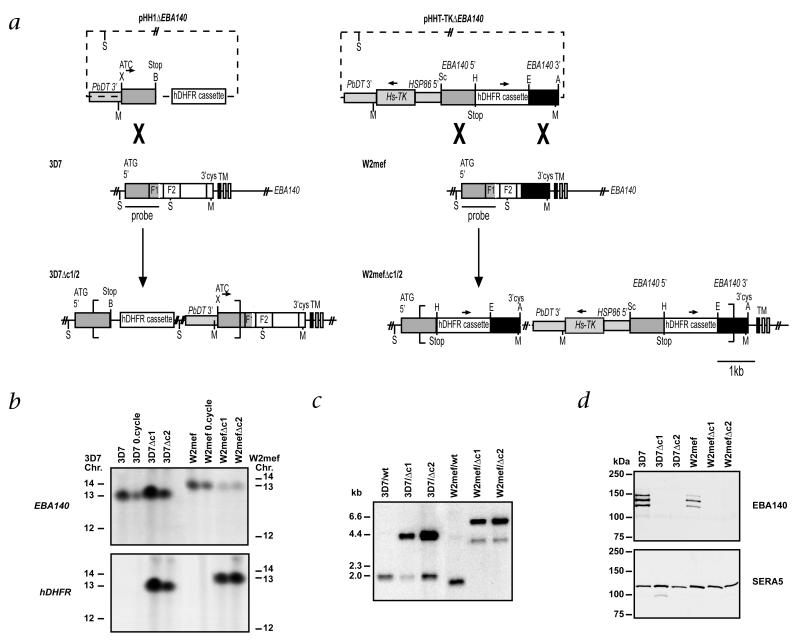
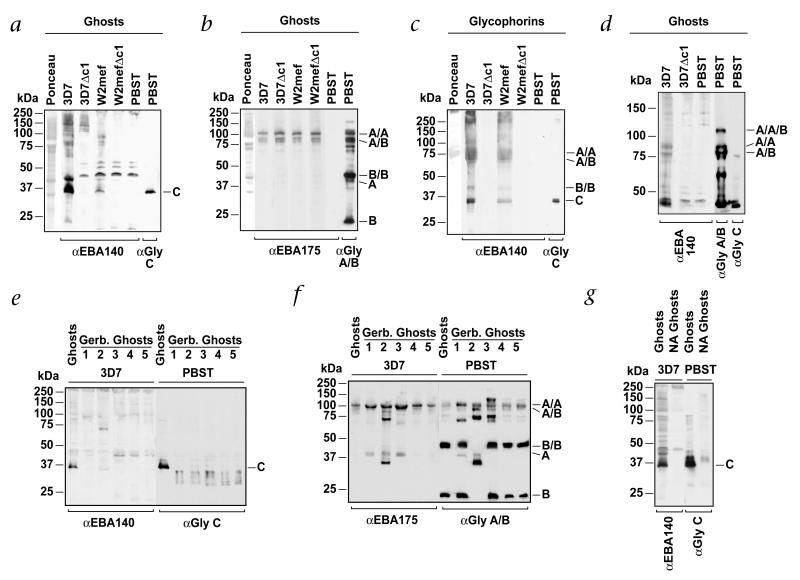
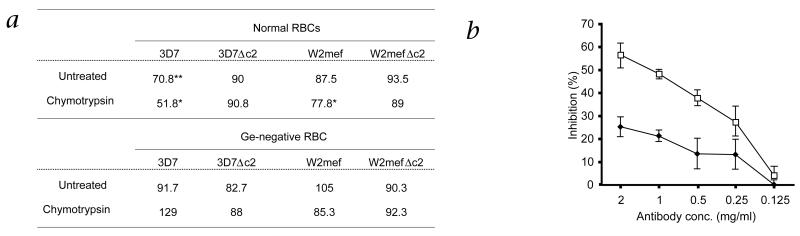
Geographic overlap between malaria and the occurrence of mutant hemoglobin and erythrocyte surface proteins has indicated that polymorphisms in human genes have been selected by severe malaria. Deletion of exon 3 in the glycophorin C gene (called GYPCDeltaex3 here) has been found in Melanesians; this alteration changes the serologic phenotype of the Gerbich (Ge) blood group system, resulting in Ge negativity. The GYPCDeltaex3 allele reaches a high frequency (46.5%) in coastal areas of Papua New Guinea where malaria is hyperendemic. The Plasmodium falciparum erythrocyte-binding antigen 140 (EBA140, also known as BAEBL) binds with high affinity to the surface of human erythrocytes. Here we show that the receptor for EBA140 is glycophorin C (GYPC) and that this interaction mediates a principal P. falciparum invasion pathway into human erythrocytes. EBA140 does not bind to GYPC in Ge-negative erythrocytes, nor can P. falciparum invade such cells using this invasion pathway. This provides compelling evidence that Ge negativity has arisen in Melanesian populations through natural selection by severe malaria.
Figures
References
-
- Haldane JBS. The rate of mutation in human genes. In: bonnier GA, editor. Proceedings of the VII International Congress on Genetics. 1949. pp. 267–273.
-
- Miller LH, Good MF, Milon G. Malaria pathogenesis. Science. 1994;264:1878–1883. - PubMed
-
- Booth PB, McLoughlin K. The Gerbich blood group system, especially in Melanesians. Vox Sang. 1972;22:73–84. - PubMed
-
- Serjeantson SW, White BS, Bhatia K, Trent RJ. A 3.5 kb deletion in the gly-cophorin C gene accounts for the Gerbich-negative blood group in Melanesians. Immunol. Cell Biol. 1994;72:23–27. - PubMed
-
- Patel SS, et al. The association of the glycophorin C exon 3 deletion with ovalocytosis and malaria susceptibility in the Wosera, Papua New Guinea. Blood. 2001;98:3489–3491. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
