

Lineage-specific requirements of beta-catenin in neural crest development
- PMID: 12473692
- PMCID: PMC2173383
- DOI: 10.1083/jcb.200209039
Lineage-specific requirements of beta-catenin in neural crest development
Abstract
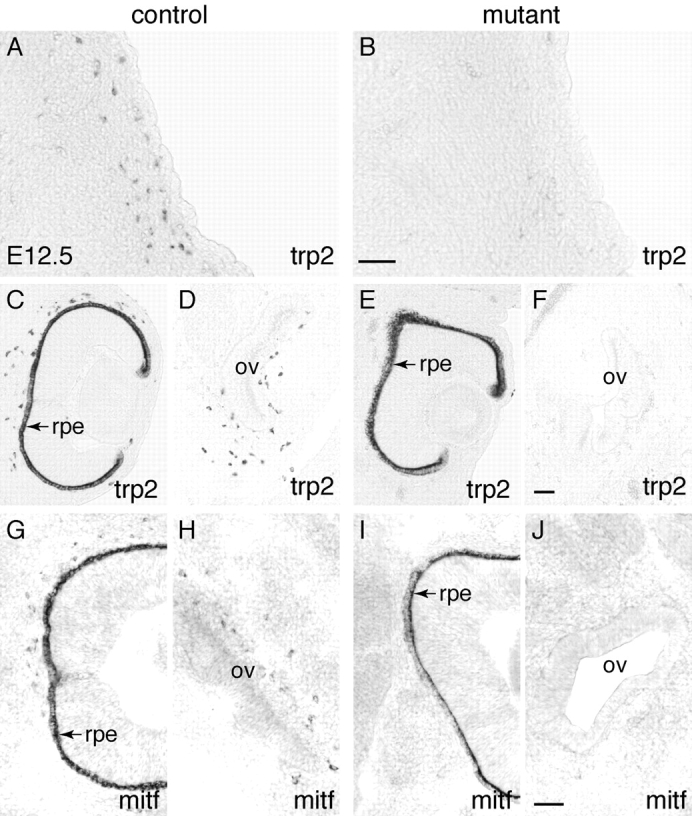
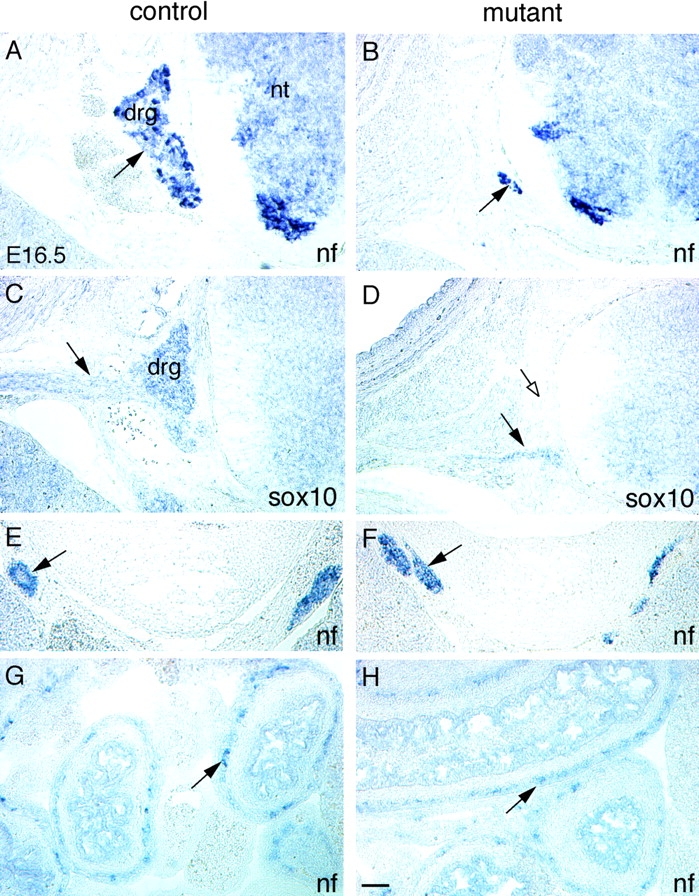
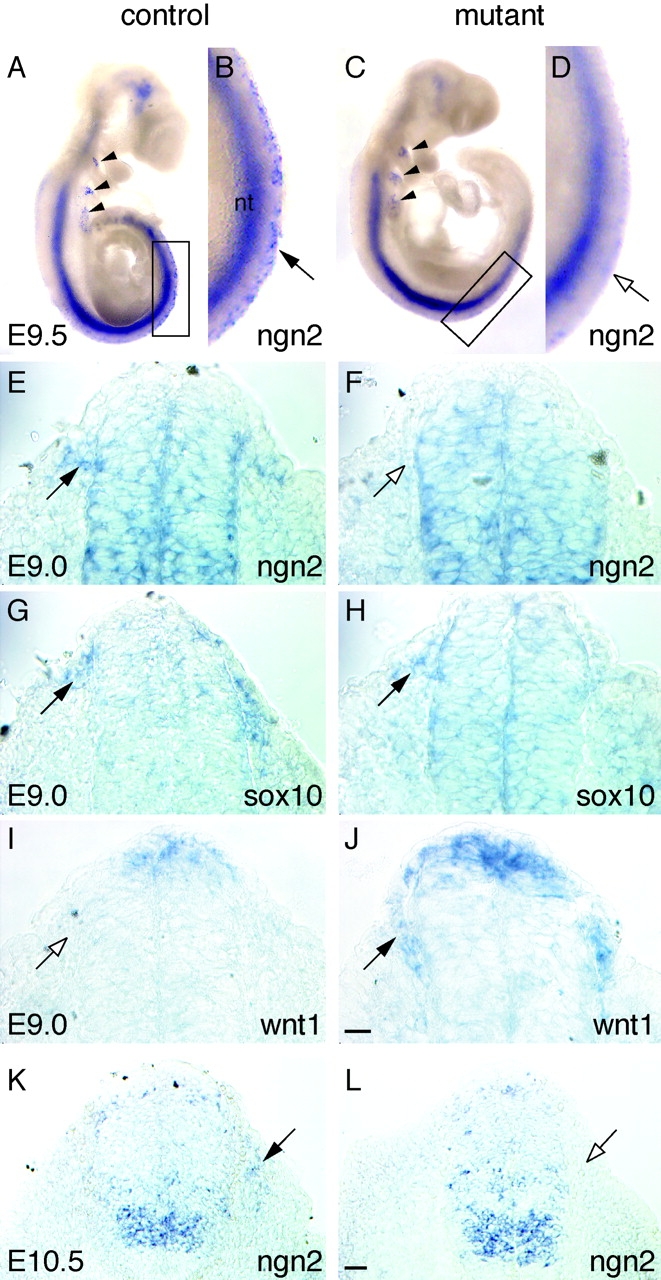
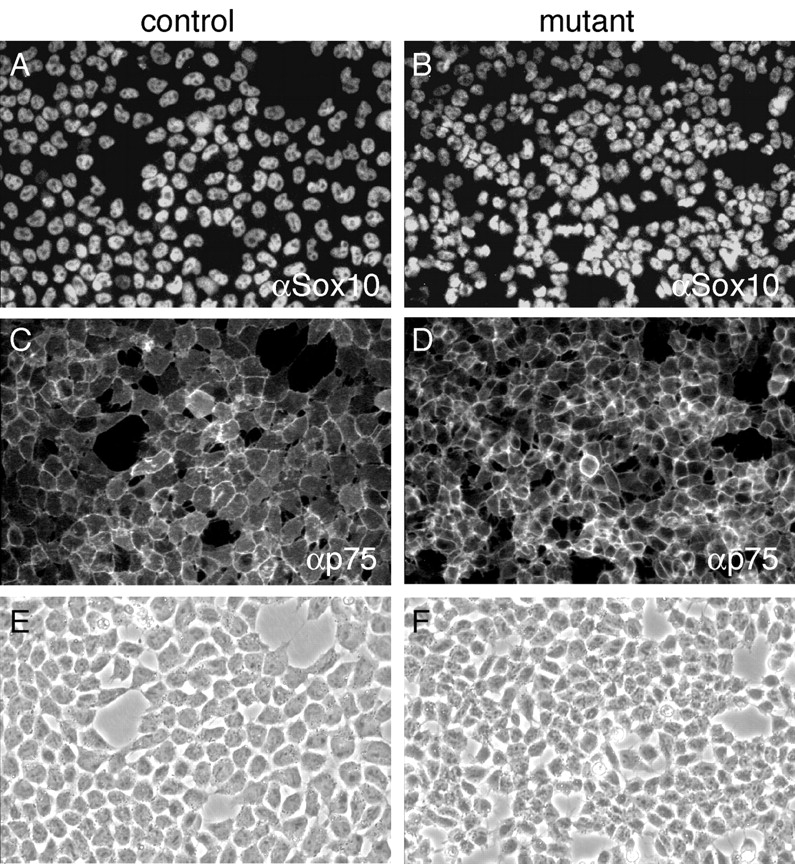
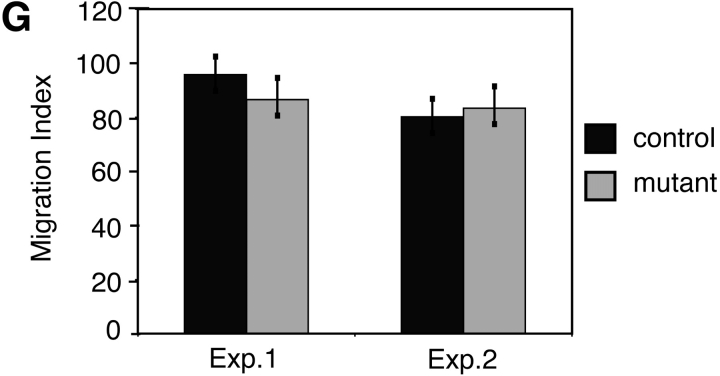
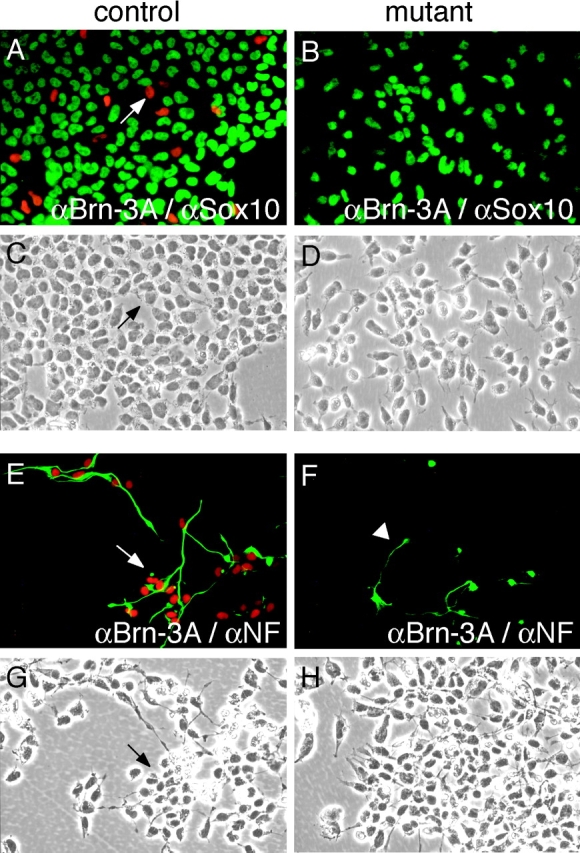
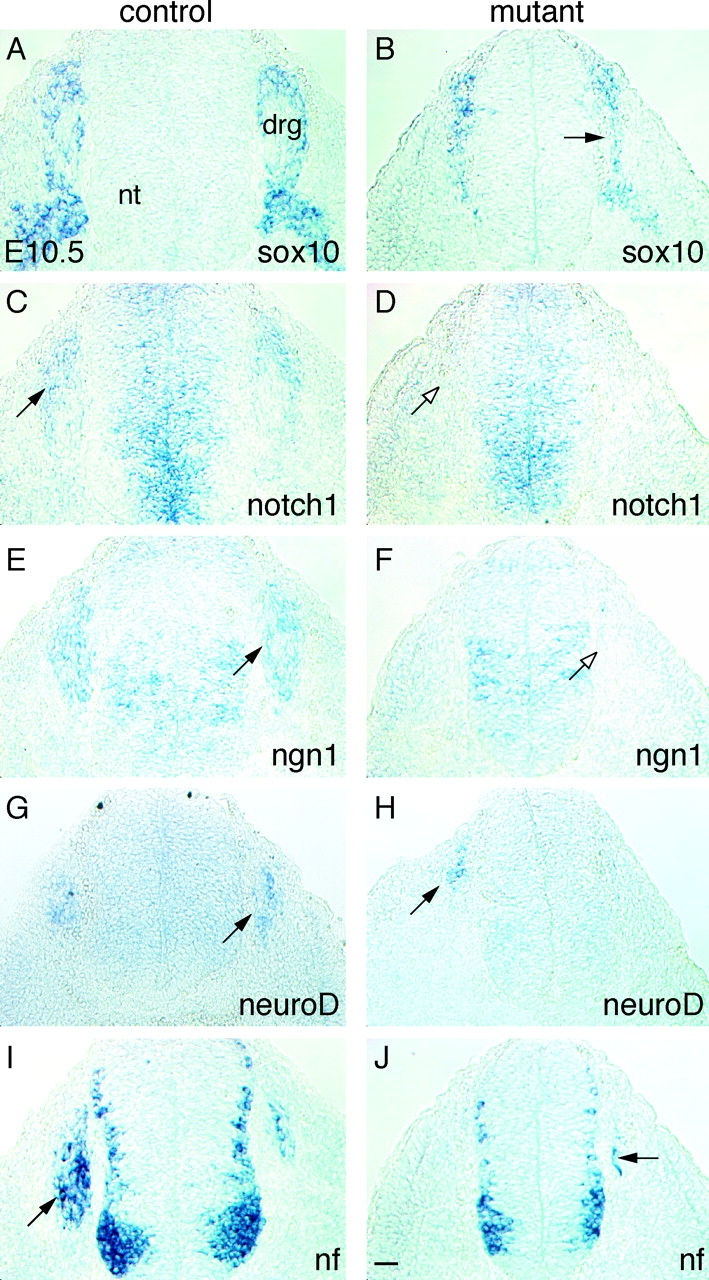
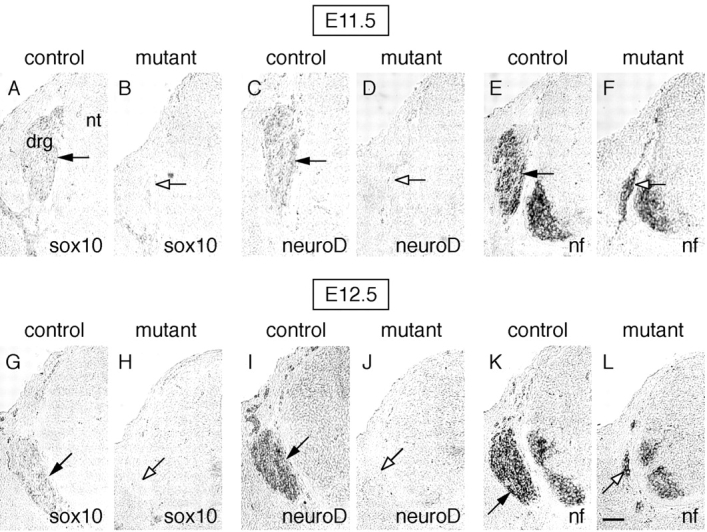
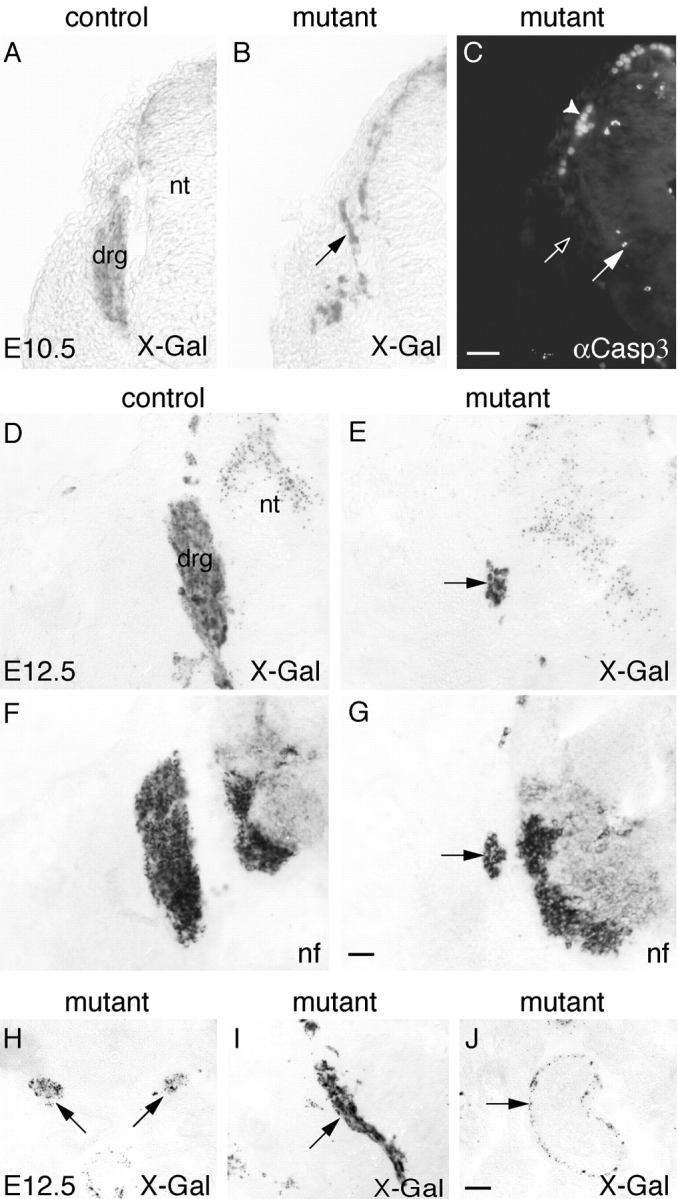
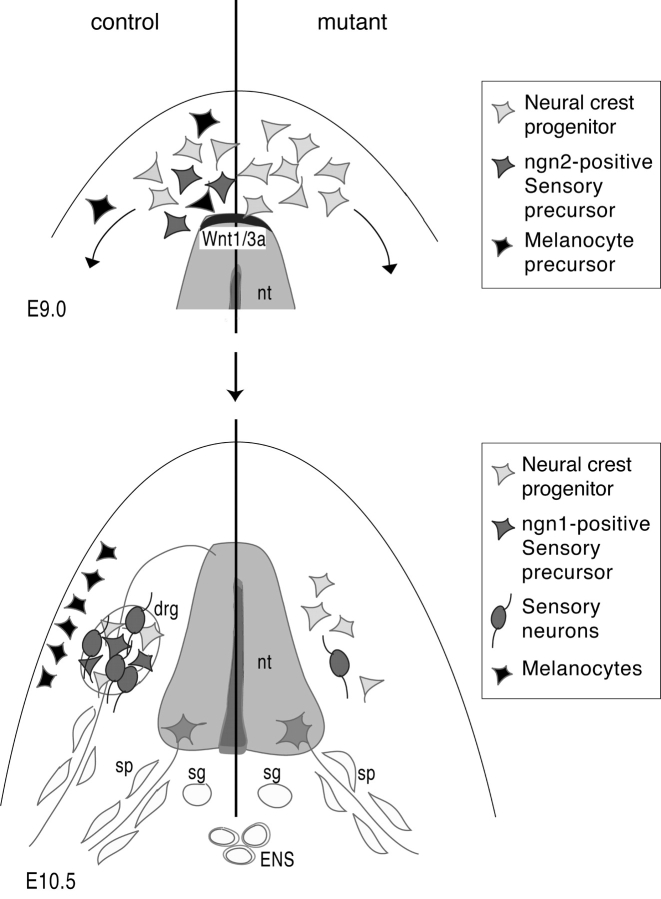
Beta-catenin plays a pivotal role in cadherin-mediated cell adhesion. Moreover, it is a downstream signaling component of Wnt that controls multiple developmental processes such as cell proliferation, apoptosis, and fate decisions. To study the role of beta-catenin in neural crest development, we used the Cre/loxP system to ablate beta-catenin specifically in neural crest stem cells. Although several neural crest-derived structures develop normally, mutant animals lack melanocytes and dorsal root ganglia (DRG). In vivo and in vitro analyses revealed that mutant neural crest cells emigrate but fail to generate an early wave of sensory neurogenesis that is normally marked by the transcription factor neurogenin (ngn) 2. This indicates a role of beta-catenin in premigratory or early migratory neural crest and points to heterogeneity of neural crest cells at the earliest stages of crest development. In addition, migratory neural crest cells lateral to the neural tube do not aggregate to form DRG and are unable to produce a later wave of sensory neurogenesis usually marked by the transcription factor ngn1. We propose that the requirement of beta-catenin for the specification of melanocytes and sensory neuronal lineages reflects roles of beta-catenin both in Wnt signaling and in mediating cell-cell interactions.
Figures
References
-
- Anderson, D.J. 1999. Lineages and transcription factors in the specification of vertebrate primary sensory neurons. Curr. Opin. Neurobiol. 9:517–524. - PubMed
-
- Anderson, D.J., A. Groves, L. Lo, Q. Ma, M. Rao, N.M. Shah, and L. Sommer. 1997. Cell lineage determination and the control of neuronal identity in the neural crest. Cold Spring Harb. Symp. Quant. Biol. 62:493–504. - PubMed
-
- Borchers, A., R. David, and D. Wedlich. 2001. Xenopus cadherin-11 restrains cranial neural crest migration and influences neural crest specification. Development. 128:3049–3060. - PubMed
-
- Brault, V., R. Moore, S. Kutsch, M. Ishibashi, D.H. Rowitch, A.P. McMahon, L. Sommer, O. Boussida, and R. Kemler. 2001. Inactivation of the β-catenin gene by Wnt1-Cre-mediated deletion results in dramatic brain malformation and failure of craniofacial development. Development. 128:1253–1264. - PubMed
-
- Cadigan, K.M., and R. Nusse. 1997. Wnt signaling: a common theme in animal development. Genes Dev. 11:3286–3305. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
