

Direct inhibition by nitric oxide of the transcriptional ferric uptake regulation protein via nitrosylation of the iron
- PMID: 12475930
- PMCID: PMC139193
- DOI: 10.1073/pnas.252591299
Direct inhibition by nitric oxide of the transcriptional ferric uptake regulation protein via nitrosylation of the iron
Abstract
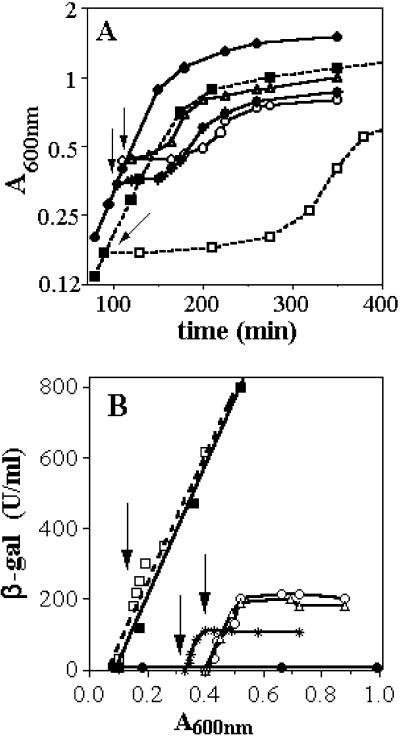
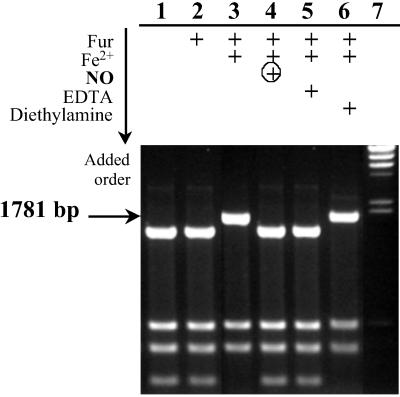
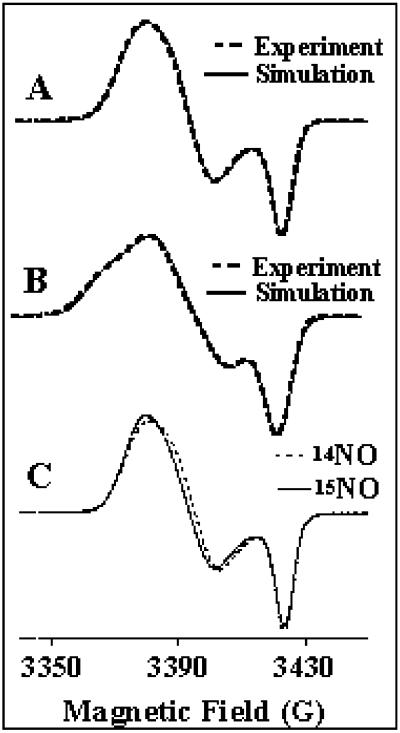
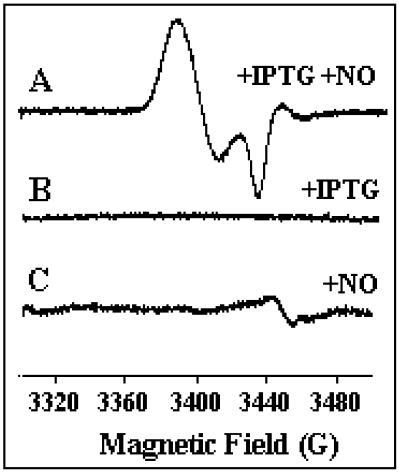
Ferric uptake regulation protein (Fur) is a bacterial global regulator that uses iron as a cofactor to bind to specific DNA sequences. The function of Fur is not limited to iron homeostasis. A wide variety of genes involved in various mechanisms such as oxidative and acid stresses are under Fur control. Flavohemoglobin (Hmp) is an NO-detoxifying enzyme induced by NO and nitrosothiol compounds. Fur recently was found to regulate hmp in Salmonella typhimurium, and in Escherichia coli, the iron-chelating agent 2,2'-dipyridyl induces hmp expression. We now establish direct inhibition of E. coli Fur activity by NO. By using chromosomal Fur-regulated lacZ reporter fusion in E. coli, Fur activity is switched off by NO at micromolar concentration. In vitro Fur DNA-binding activity, as measured by protection of restriction site in aerobactin promoter, is directly sensitive to NO. NO reacts with Fe(II) in purified FeFur protein to form a S = 12 low-spin FeFur-NO complex with a g = 2.03 EPR signal. Appearance of the same EPR signal in NO-treated cells links nitrosylation of the iron with Fur inhibition. The nitrosylated Fur protein is still a dimer and is stable in anaerobiosis but slowly decays in air. This inhibition probably arises from a conformational switch, leading to an inactive dimeric protein. These data establish a link between control of iron metabolism and the response to NO effects.
Figures
References
-
- Neilands J. B. (1972) Struct. Bonding (Berlin) 11, 145-170.
-
- Hibbs J. B., Taintor, R. R., Vavrin, Z. & Rachlin, E. M. (1988) Biochem. Biophys. Res. Commun. 157, 87-94. - PubMed
-
- Marletta M. A., Yoon, P. S., Iyengar, R., Leaf, C. D. & Wishnok, J. S. (1988) Biochemistry 27, 8706-8711. - PubMed
-
- Sari M.-A., Moali, C., Boucher, J. L., Jaouen, M. & Mansuy, D. (1998) Biochem. Biophys. Res. Commun. 250, 364-368. - PubMed
-
- Adak S., Aulak, K. S. & Stuehr, D. J. (2002) J. Biol. Chem. 277, 16167-16171. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
