

Regulation of IkappaBbeta expression in testis
- PMID: 12475944
- PMCID: PMC138625
- DOI: 10.1091/mbc.01-07-0373
Regulation of IkappaBbeta expression in testis
Abstract
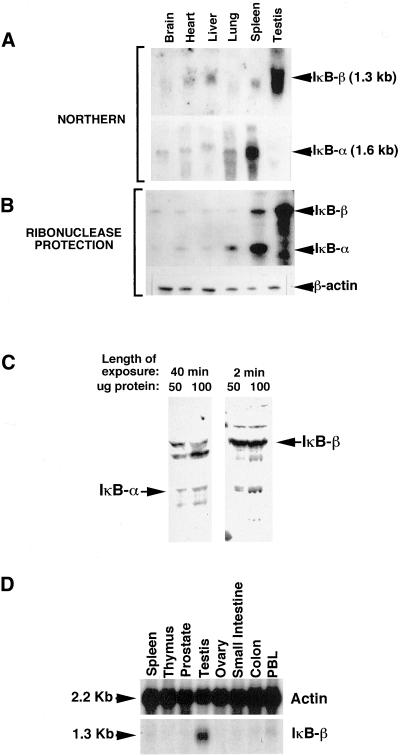
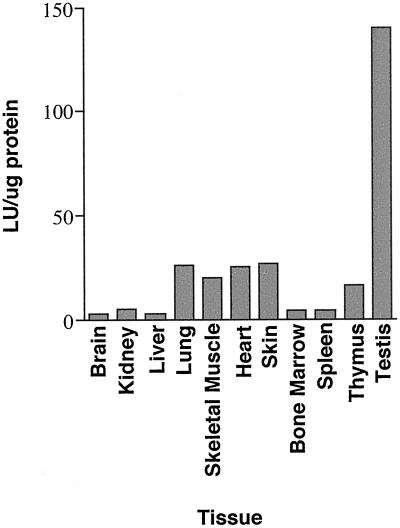
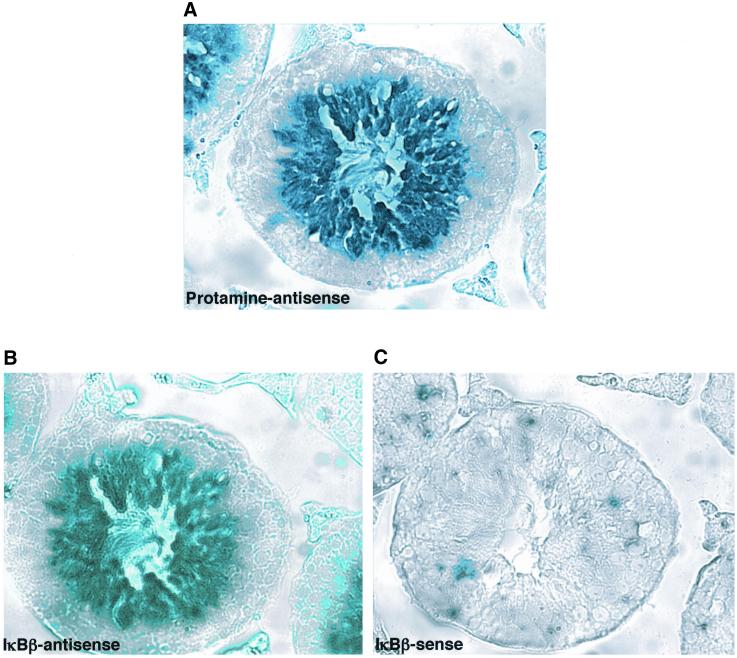
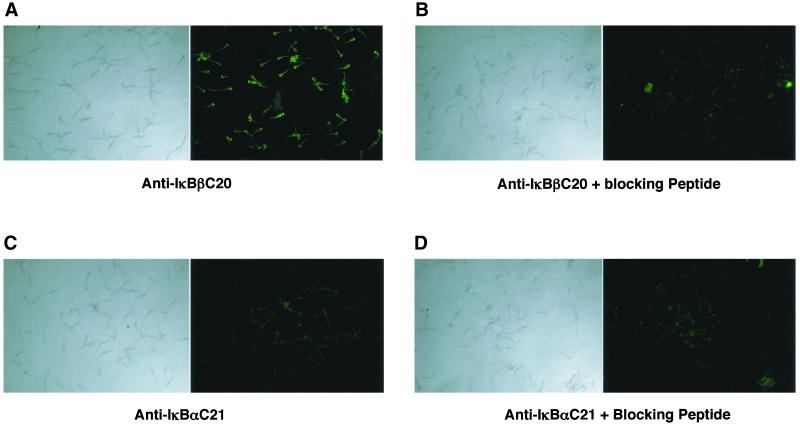
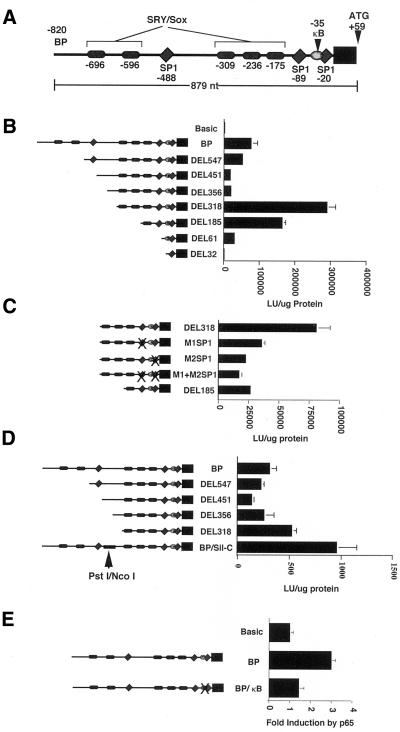
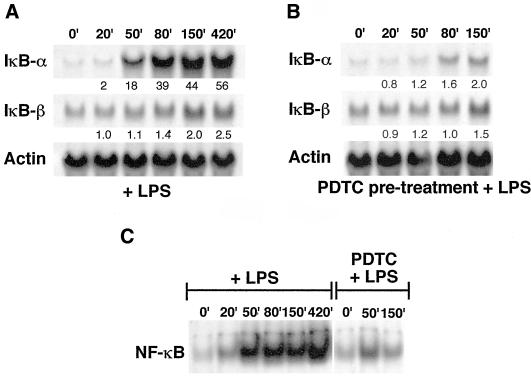
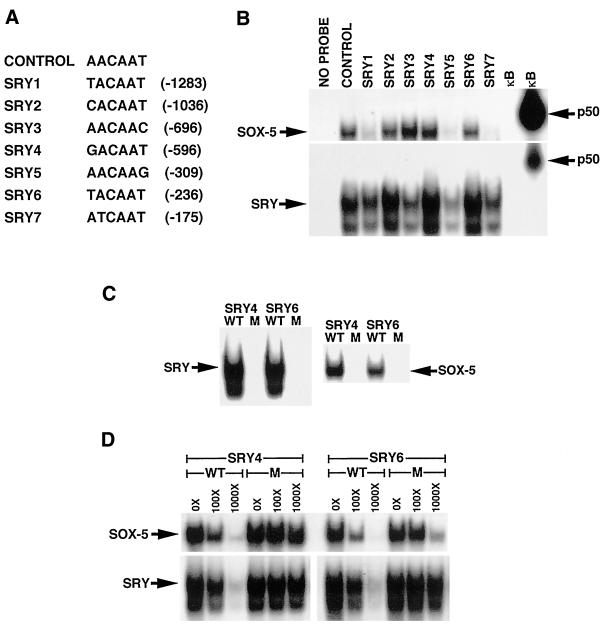
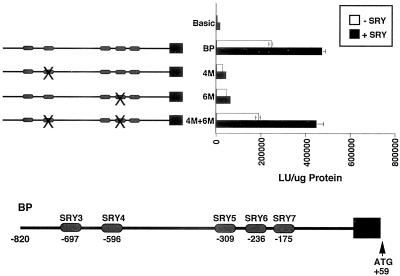
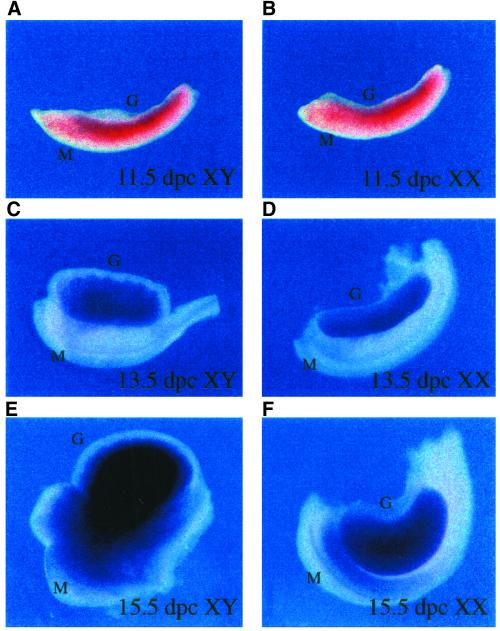
IkappaBalpha and IkappaBbeta are regulators of the nuclear factor-kappaB (NF-kappaB) transcription factor family. Both IkappaBs bind to the same NF-kappaB dimers and are widely expressed in different cells and tissues. To better understand how these two IkappaB isoforms differ biologically, we have characterized the expression of IkappaBbeta in testis, a tissue in which IkappaBalpha is only minimally expressed. We have found that IkappaBbeta expression is localized within the haploid spermatid stages of spermatogenesis and follows the expression of nuclear NF-kappaB. IkappaBbeta expression in haploid spermatids is likely regulated by Sox family proteins, members of which are also expressed within spermatids. We have shown that both SRY and Sox-5 can bind to multiple Sox binding sites found within the IkappaBbeta promoter and can enhance transcription of a reporter gene in transient transfection assays. We also demonstrate that IkappaBbeta mRNA is strongly expressed in developing male gonads. These results therefore suggest that IkappaBbeta may be a novel target for transcription factors of the HMG-box SRY/Sox family and imply a potential role for NF-kappaB/IkappaBbeta in spermatogenesis.
Figures
References
-
- Bartke A. Apoptosis of male germ cells, a generalized or a cell type-specific phenomenon? Endocrinology. 1995;136:3–4. - PubMed
-
- Beg AA, Baldwin AS., Jr The I kappa B proteins: multifunctional regulators of Rel/NF-kappa B transcription factors. Genes Dev. 1993;7:2064–2070. - PubMed
-
- Beg AA, Sha WC, Bronson RT, Baltimore D. Constitutive NF-kappa B activation, enhanced granulopoiesis, and neonatal lethality in I kappa B alpha-deficient mice. Genes Dev. 1995;9:2736–2746. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
