

Second-generation rabies virus-based vaccine vectors expressing human immunodeficiency virus type 1 gag have greatly reduced pathogenicity but are highly immunogenic
- PMID: 12477829
- PMCID: PMC140592
- DOI: 10.1128/jvi.77.1.237-244.2003
Second-generation rabies virus-based vaccine vectors expressing human immunodeficiency virus type 1 gag have greatly reduced pathogenicity but are highly immunogenic
Abstract
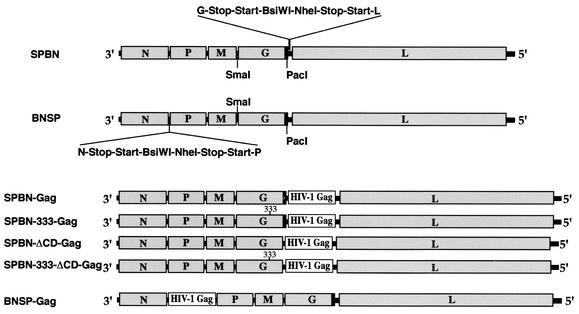
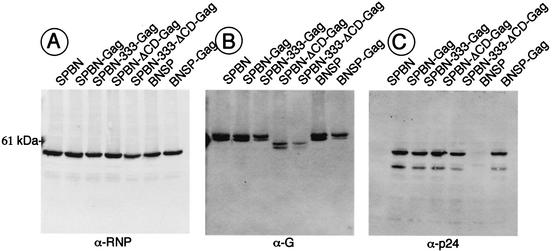
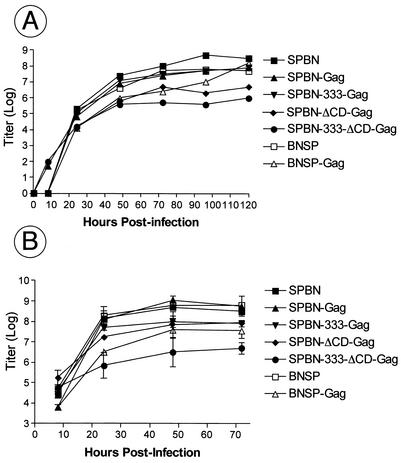
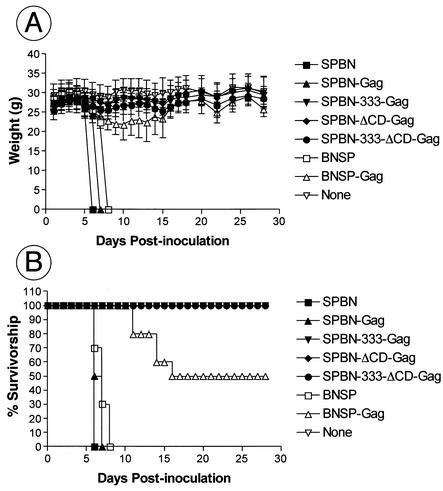
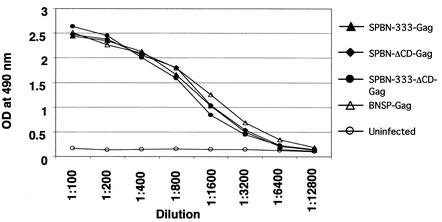
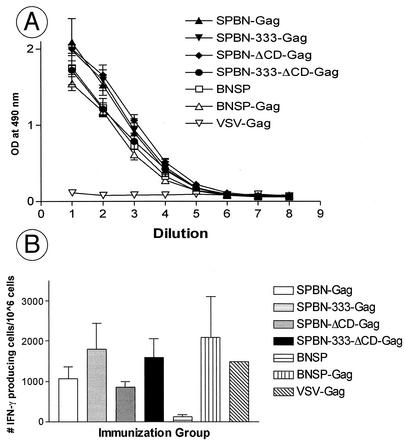
Rabies virus (RV) vaccine strain-based vectors show great promise as vaccines against other viral diseases such as human immunodeficiency virus type 1 (HIV-1) infection and hepatitis C, but a low residual pathogenicity remains a concern for their use. Here we describe several highly attenuated second-generation RV-based vaccine vehicles expressing HIV-1 Gag. For this approach, we modified the previously described RV vaccine vector SPBN by replacing the arginine at position 333 (R333) within the RV glycoprotein (G) with glutamic acid (E333), deleting 43 amino acids of the RV G cytoplasmic domain (CD), or combining the R333 exchange and the CD deletion. In addition, we constructed a new RV vector that expresses HIV-1 Gag from an RV transcription unit upstream of the RV phosphoprotein gene (BNSP-Gag) instead of upstream of the G gene. As expected and as demonstrated for SPBN-Gag, all vaccine vehicles were apathogenic after peripheral administration. However, the new, second-generation vaccine vectors containing modifications in the RV G were also apathogenic after intracranial infection with 10(5) infectious particles, and BNSP-Gag produced a 50%-reduced mortality in mice. Of note, the observed attenuation of pathogenicity did not result in either the attenuation of the humoral response against the RV G or the previously observed robust cellular response against HIV-1 Gag. These findings demonstrate that very safe and highly effective RV-based vaccines can be constructed and further emphasize their potential utility as efficacious antiviral vaccines.
Figures
References
-
- Buchholz, U. J., S. Finke, and K. K. Conzelmann. 1999. Generation of bovine respiratory syncytial virus (BRSV) from cDNA: BRSV NS2 is not essential for virus replication in tissue culture, and the human RSV leader region acts as a functional BRSV genome promoter. J. Virol. 73:251-259. - PMC - PubMed
-
- Conzelmann, K. K., J. H. Cox, L. G. Schneider, and H. J. Thiel. 1990. Molecular cloning and complete nucleotide sequence of the attenuated rabies virus SAD B19. Virology 175:485-499. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
