

Circadian clock protein KaiC forms ATP-dependent hexameric rings and binds DNA
- PMID: 12477935
- PMCID: PMC139293
- DOI: 10.1073/pnas.262578499
Circadian clock protein KaiC forms ATP-dependent hexameric rings and binds DNA
Abstract
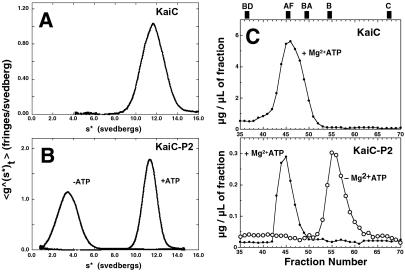
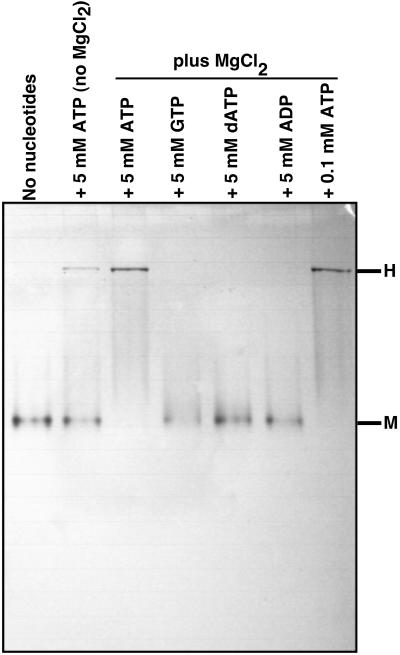
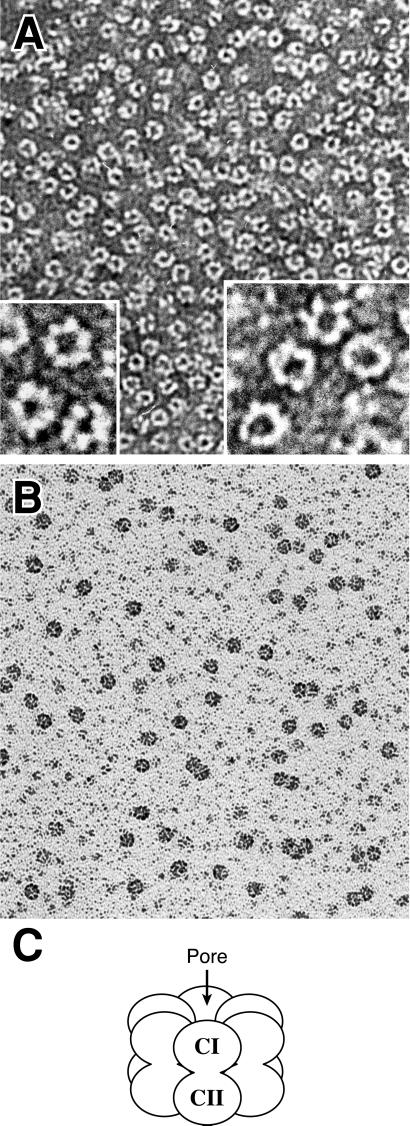
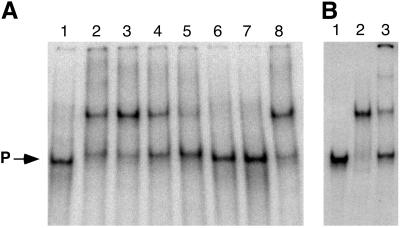
KaiC from Synechococcus elongatus PCC 7942 (KaiC) is an essential circadian clock protein in cyanobacteria. Previous sequence analyses suggested its inclusion in the RecADnaB superfamily. A characteristic of the proteins of this superfamily is that they form homohexameric complexes that bind DNA. We show here that KaiC also forms ring complexes with a central pore that can be visualized by electron microscopy. A combination of analytical ultracentrifugation and chromatographic analyses demonstrates that these complexes are hexameric. The association of KaiC molecules into hexamers depends on the presence of ATP. The KaiC sequence does not include the obvious DNA-binding motifs found in RecA or DnaB. Nevertheless, KaiC binds forked DNA substrates. These data support the inclusion of KaiC into the RecADnaB superfamily and have important implications for enzymatic activity of KaiC in the circadian clock mechanism that regulates global changes in gene expression patterns.
Figures

Similar articles
-
ATP-induced hexameric ring structure of the cyanobacterial circadian clock protein KaiC.Genes Cells. 2003 Mar;8(3):287-96. doi: 10.1046/j.1365-2443.2003.00633.x. Genes Cells. 2003. PMID: 12622725
-
KaiC from a cyanobacterium Gloeocapsa sp. PCC 7428 retains functional and structural properties required as the core of circadian clock system.Int J Biol Macromol. 2019 Jun 15;131:67-73. doi: 10.1016/j.ijbiomac.2019.03.051. Epub 2019 Mar 8. Int J Biol Macromol. 2019. PMID: 30857964
-
Role of KaiC phosphorylation in the circadian clock system of Synechococcus elongatus PCC 7942.Proc Natl Acad Sci U S A. 2004 Sep 21;101(38):13927-32. doi: 10.1073/pnas.0403906101. Epub 2004 Sep 3. Proc Natl Acad Sci U S A. 2004. PMID: 15347812 Free PMC article.
-
A cyanobacterial circadian clock based on the Kai oscillator.Cold Spring Harb Symp Quant Biol. 2007;72:47-55. doi: 10.1101/sqb.2007.72.029. Cold Spring Harb Symp Quant Biol. 2007. PMID: 18419262 Review.
-
Diversity of KaiC-based timing systems in marine Cyanobacteria.Mar Genomics. 2014 Apr;14:3-16. doi: 10.1016/j.margen.2013.12.006. Epub 2014 Jan 3. Mar Genomics. 2014. PMID: 24388874 Review.
Cited by
-
Synechocystis KaiC3 Displays Temperature- and KaiB-Dependent ATPase Activity and Is Important for Growth in Darkness.J Bacteriol. 2020 Jan 29;202(4):e00478-19. doi: 10.1128/JB.00478-19. Print 2020 Jan 29. J Bacteriol. 2020. PMID: 31767776 Free PMC article.
-
Temporal and spatial oscillations in bacteria.Nat Rev Microbiol. 2011 Aug 15;9(8):565-77. doi: 10.1038/nrmicro2612. Nat Rev Microbiol. 2011. PMID: 21760621 Review.
-
Tracking and visualizing the circadian ticking of the cyanobacterial clock protein KaiC in solution.EMBO J. 2011 Jan 5;30(1):68-78. doi: 10.1038/emboj.2010.298. Epub 2010 Nov 26. EMBO J. 2011. PMID: 21113137 Free PMC article.
-
Intricate protein-protein interactions in the cyanobacterial circadian clock.J Biol Chem. 2014 Aug 1;289(31):21267-75. doi: 10.1074/jbc.R114.579607. Epub 2014 Jun 16. J Biol Chem. 2014. PMID: 24936066 Free PMC article. Review.
-
Loop-loop interactions regulate KaiA-stimulated KaiC phosphorylation in the cyanobacterial KaiABC circadian clock.Biochemistry. 2013 Feb 19;52(7):1208-20. doi: 10.1021/bi301691a. Epub 2013 Feb 7. Biochemistry. 2013. PMID: 23351065 Free PMC article.
References
-
- Johnson C. H., Golden, S. S., Ishiura, M. & Kondo, T. (1996) Mol. Microbiol. 21, 5-11. - PubMed
-
- Johnson C. H. (2001) Annu. Rev. Physiol. 63, 695-728. - PubMed
-
- Mori T. & Johnson, C. H. (2001) Semin. Cell Dev. Biol. 12, 271-278. - PubMed
-
- Kondo T., Tsinoremas, N. F., Golden, S. S., Johnson, C. H., Kutsuna, S. & Ishiura, M. (1994) Science 266, 1233-1236. - PubMed
-
- Ishiura M., Kutsuna, S., Aoki, S., Iwasaki, H., Andersson, C. R., Tanabe, A., Golden, S. S., Johnson, C. H. & Kondo, T. (1998) Science 281, 1519-1523. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
