

Smooth muscle membrane potential modulates endothelium-dependent relaxation of rat basilar artery via myo-endothelial gap junctions
- PMID: 12482900
- PMCID: PMC2290719
- DOI: 10.1113/jphysiol.2002.031823
Smooth muscle membrane potential modulates endothelium-dependent relaxation of rat basilar artery via myo-endothelial gap junctions
Abstract
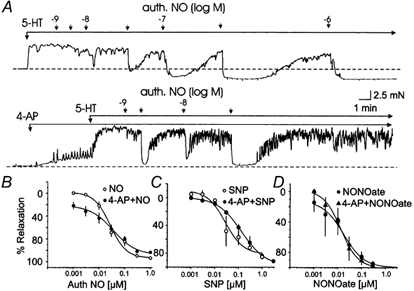
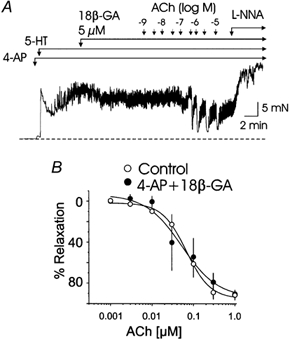
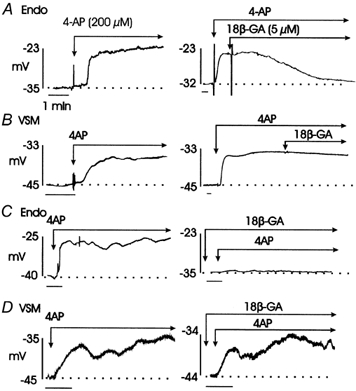
The release of endothelium-derived relaxing factors, such as nitric oxide (NO), is dependent on an increase in intracellular calcium levels ([Ca(2+)](i)) within endothelial cells. Endothelial cell membrane potential plays a critical role in the regulation of [Ca(2+)](i) in that calcium influx from the extracellular space is dependent on membrane hyperpolarization. In this study, the effect of inhibition of vascular smooth muscle delayed rectifier K(+) (K(DR)) channels by 4-aminopyridine (4-AP) on endothelium-dependent relaxation of rat basilar artery to acetylcholine (ACh) was assessed. ACh-evoked endothelium-dependent relaxations were inhibited by N-(Omega)-nitro-L-arginine (L-NNA) or 1H-[1,2,4]oxadiazolo[4,3-a]quinoxalin-1-one (ODQ), confirming a role for NO and guanylyl cyclase. 4-AP (300 microM) also suppressed ACh-induced relaxation, with the maximal response reduced from approximately 92 to approximately 33 % (n = 11; P < 0.01). However, relaxations in response to exogenous NO, applied in the form of authentic NO, sodium nitroprusside or diethylamineNONOate (DEANONOate), were not affected by 4-AP treatment (n = 3-11). These data are not consistent with the view that 4-AP-sensitive K(DR) channels are mediators of vascular hyperpolarization and relaxation in response to endothelium-derived NO. Inhibition of ACh-evoked relaxation by 4-AP was reversed by pinacidil (0.5-1 microM; n = 5) or 18beta-glycyrrhetinic acid (18betaGA; 5 microM; n = 5), indicating that depolarization and electrical coupling of the smooth muscle to the endothelium were involved. 4-AP caused depolarization of both endothelial and vascular smooth muscle cells of isolated segments of basilar artery (mean change 11 +/- 1 and 9 +/- 2 mV, respectively; n = 15). Significantly, 18betaGA almost completely prevented the depolarization of endothelial cells (n = 6), but not smooth muscle cells (n = 6) by 4-AP. ACh-induced hyperpolarization of endothelium and smooth muscle cells was also reduced by 4-AP, but this inhibition was not observed in the combined presence of 4-AP and 18betaGA. These data indicate that 4-AP can induce an indirect inhibition of endothelium-dependent relaxation in the rat basilar artery by electrical coupling of smooth muscle membrane depolarization to the endothelium via myo-endothelial gap junctions.
Figures





References
-
- Adams DJ, Barakeh J, Laskey R, Van Breeman C. Ion channels and regulation of intracellular calcium in vascular endothelial cells. FASEB Journal. 1989;3:2389–2400. - PubMed
-
- Aiello EA, Malcolm AT, Walsh MP, Cole WC. Beta-adrenoceptor activation and PKA regulate delayed rectifier K+ channels of vascular smooth muscle cells. American Journal of Physiology. 1998;275:H448–459. - PubMed
-
- Aiello EA, Walsh MP, Cole WC. Phosphorylation by protein kinase A enhances delayed rectifier K+ current in vascular smooth muscle cells. American Journal of Physiology. 1995;268:H924–934. - PubMed
-
- Akhtar S, Shamotienko O, Papakosta M, Ali F, Dolly JO. Characteristics of brain Kv1 channels tailored to mimic native counterparts by tandem linkage of alpha subunits. Implications for K+ channelopathies. Journal of Biological Chemistry. 2002;277:16376–16383. - PubMed
-
- Baron AD. Insulin resistance and vascular function. Journal of Diabetes Complications. 2002;16:92–102. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous