

Activating signal cointegrator 2 belongs to a novel steady-state complex that contains a subset of trithorax group proteins
- PMID: 12482968
- PMCID: PMC140670
- DOI: 10.1128/MCB.23.1.140-149.2003
Activating signal cointegrator 2 belongs to a novel steady-state complex that contains a subset of trithorax group proteins
Abstract
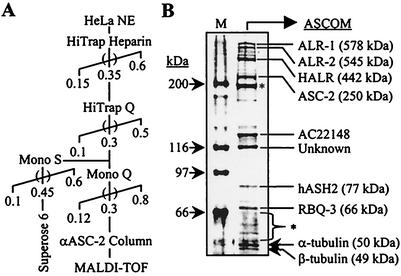
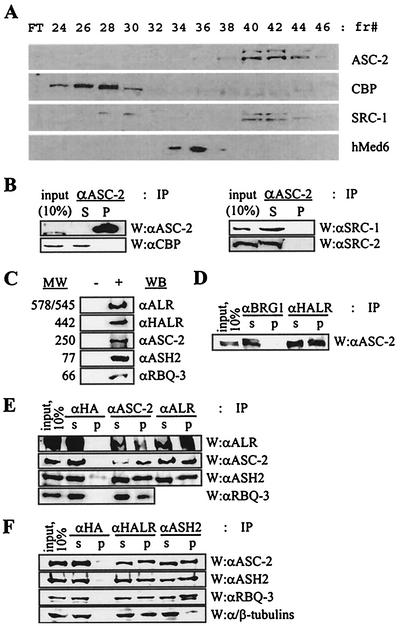
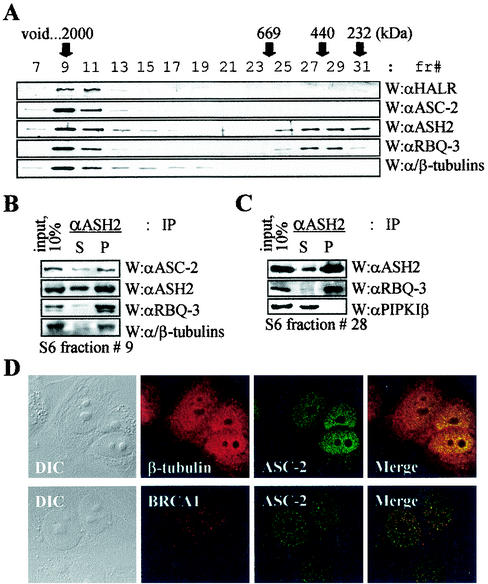
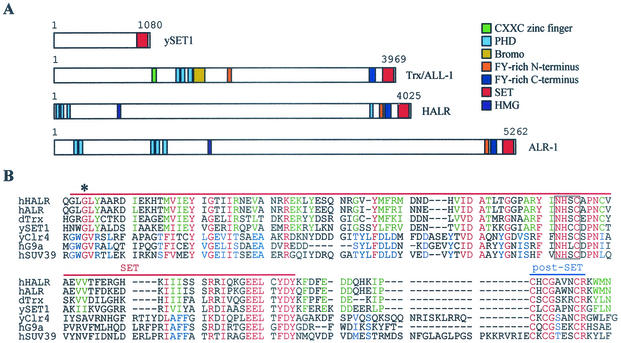
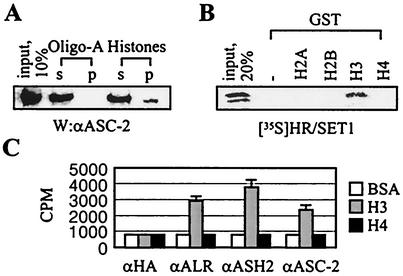
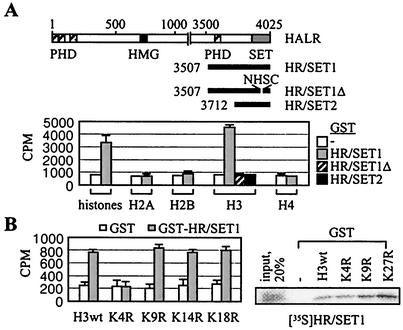
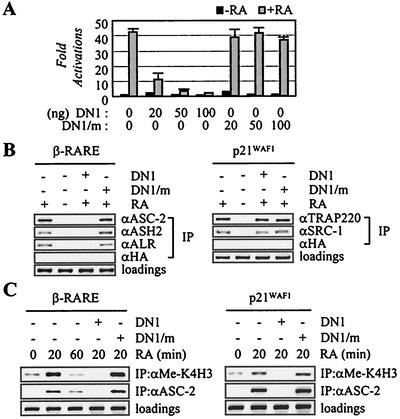
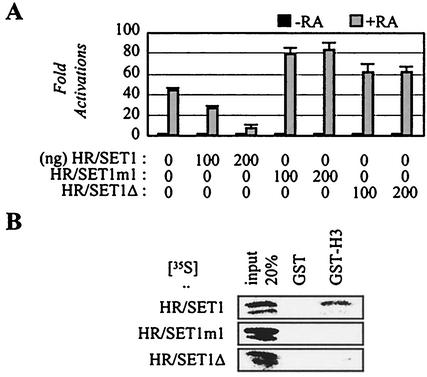
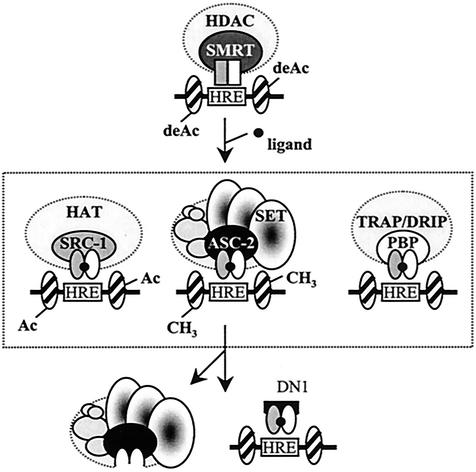
Many transcription coactivators interact with nuclear receptors in a ligand- and C-terminal transactivation function (AF2)-dependent manner. These include activating signal cointegrator 2 (ASC-2), a recently isolated transcriptional coactivator molecule, which is amplified in human cancers and stimulates transactivation by nuclear receptors and numerous other transcription factors. In this report, we show that ASC-2 belongs to a steady-state complex of approximately 2 MDa (ASC-2 complex [ASCOM]) in HeLa nuclei. ASCOM contains retinoblastoma-binding protein RBQ-3, alpha/beta-tubulins, and trithorax group proteins ALR-1, ALR-2, HALR, and ASH2. In particular, ALR-1/2 and HALR contain a highly conserved 130- to 140-amino-acid motif termed the SET domain, which was recently implicated in histone H3 lysine-specific methylation activities. Indeed, recombinant ALR-1, HALR, and immunopurified ASCOM exhibit very weak but specific H3-lysine 4 methylation activities in vitro, and transactivation by retinoic acid receptor appears to involve ligand-dependent recruitment of ASCOM and subsequent transient H3-lysine 4 methylation of the promoter region in vivo. Thus, ASCOM may represent a distinct coactivator complex of nuclear receptors. Further characterization of ASCOM will lead to a better understanding of how nuclear receptors and other transcription factors mediate transcriptional activation.
Figures
References
-
- Ayton, P. M., and M. L. Cleary. 2001. Molecular mechanisms of leukemogenesis mediated by MLL fusion proteins. Oncogene 20:5695-5707. - PubMed
-
- Bernstam, V. A., R. H. Gray, and I. A. Bernstein. 1980. Effects of microtubule-disrupting drugs on protein and RNA synthesis in Physarum polycephalum amoebae. Arch. Microbiol. 128:34-40. - PubMed
-
- Caira, F., P. Antonson, M. Pelto-Huikko, E. Treuter, and J. A. Gustafsson. 2000. Cloning and characterization of RAP250, a novel nuclear receptor coactivator. J. Biol. Chem. 275:5308-5317. - PubMed
-
- Chen, D., H. Ma, H. Hong, S. S. Koh, S. M. Huang, B. T. Schurter, D. W. Aswad, and M. R. Stallcup. 1999. Regulation of transcription by a protein methyltransferase. Science 284:2174-2177. - PubMed
-
- Cremer, T., G. Kreth, H. Koester, R. H. Fink, R. Heintzmann, M. Cremer, I. Solovei, D. Zink, and C. Cremer. 2000. Chromosome territories, interchromatin domain compartment, and nuclear matrix: an integrated view of the functional nuclear architecture. Crit. Rev. Eukaryot. Gene Expr. 10:179-212. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous