

Adaptation to Fe-deficiency requires remodeling of the photosynthetic apparatus
- PMID: 12485992
- PMCID: PMC139087
- DOI: 10.1093/emboj/cdf666
Adaptation to Fe-deficiency requires remodeling of the photosynthetic apparatus
Abstract
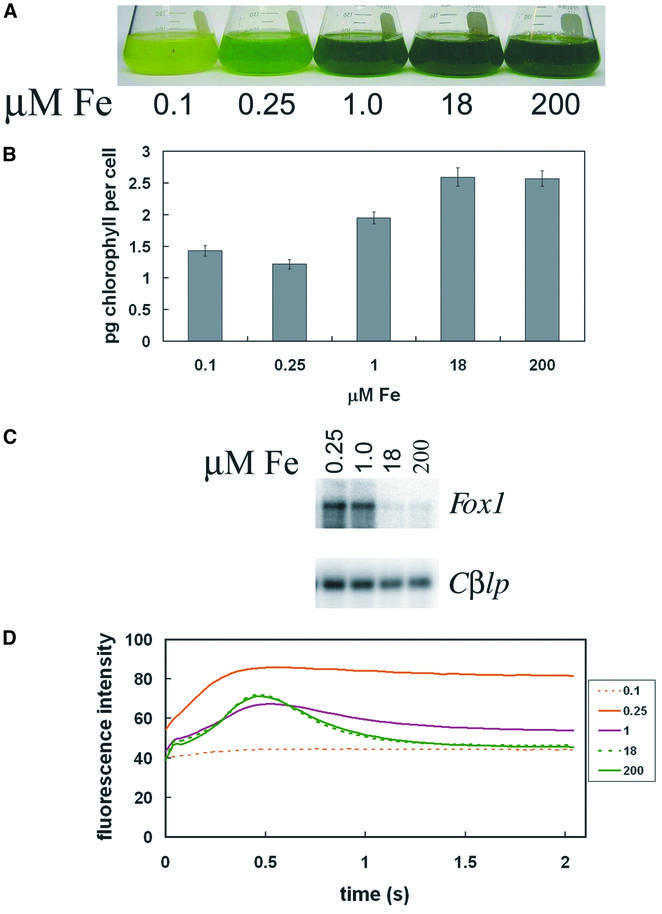
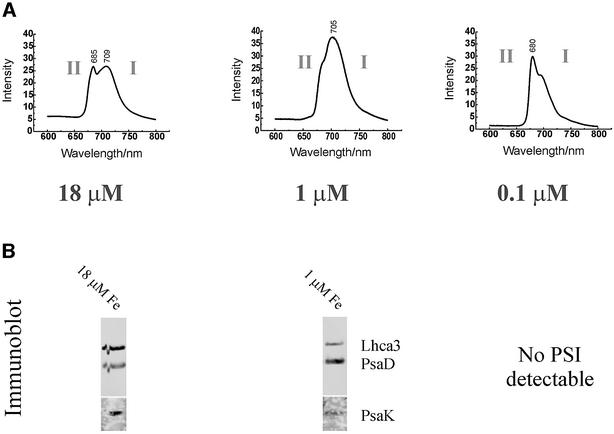
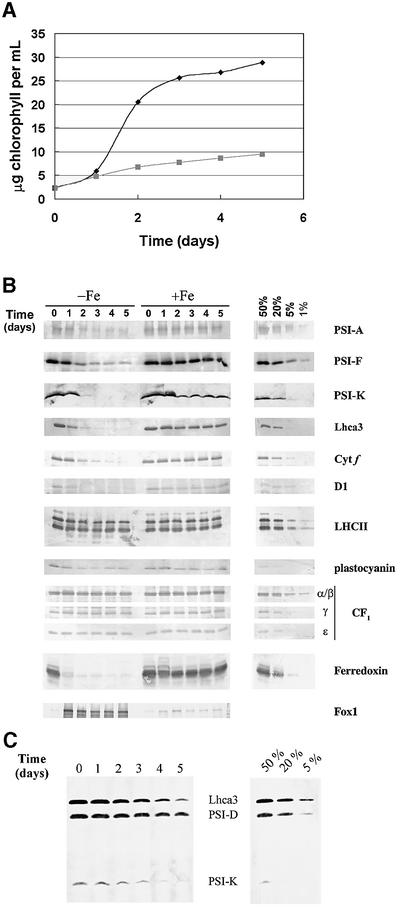
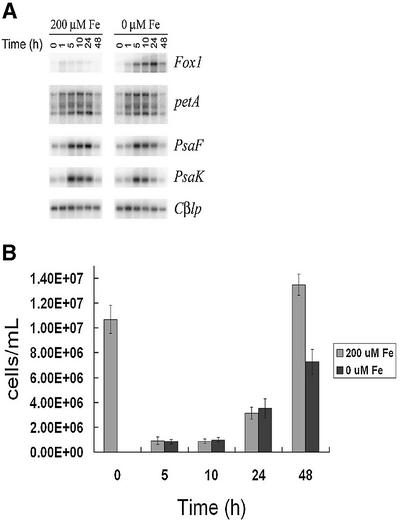
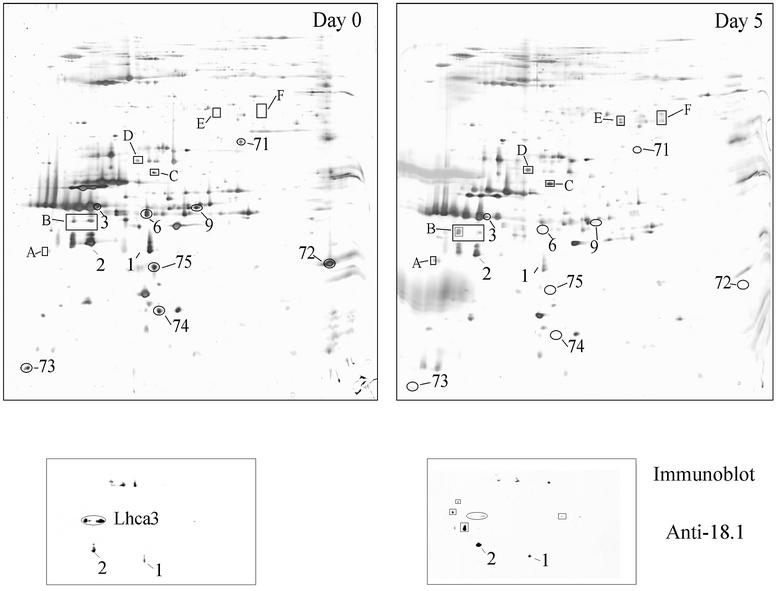
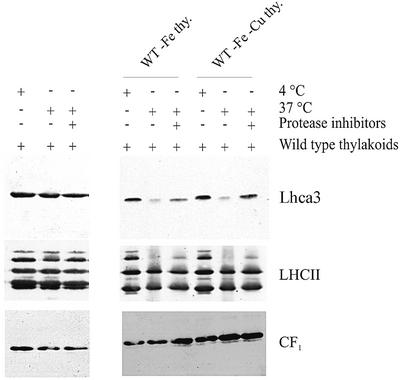
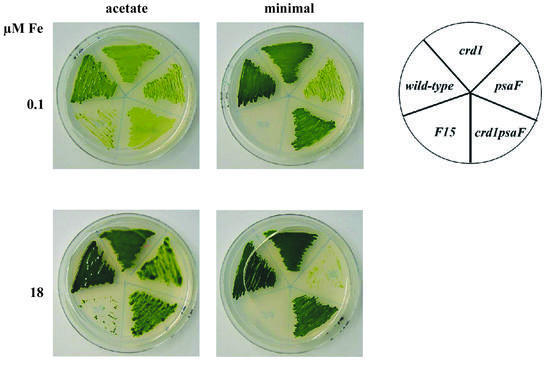
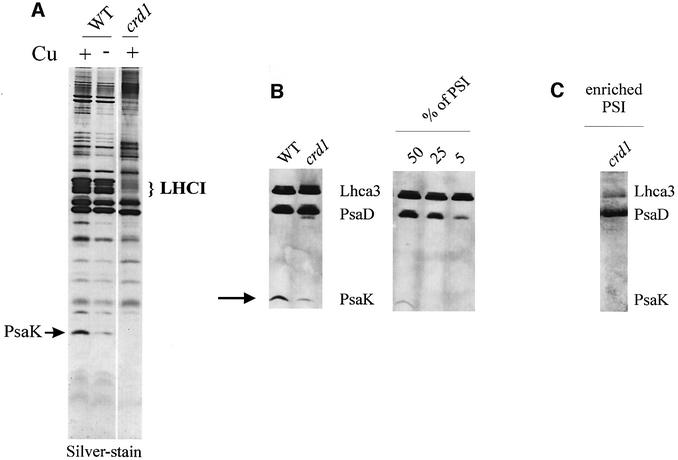
The molecular mechanisms underlying the onset of Fe-deficiency chlorosis and the maintenance of photosynthetic function in chlorotic chloroplasts are relevant to global photosynthetic productivity. We describe a series of graded responses of the photosynthetic apparatus to Fe-deficiency, including a novel response that occurs prior to the onset of chlorosis, namely the disconnection of the LHCI antenna from photosystem I (PSI). We propose that disconnection is mediated by a change in the physical properties of PSI-K in PSI in response to a change in plastid Fe content, which is sensed through the occupancy, and hence activity, of the Fe-containing active site in Crd1. We show further that progression of the response involves remodeling of the antenna complexes-specific degradation of existing proteins coupled to the synthesis of new ones, and establishment of a new steady state with decreased stoichiometry of electron transfer complexes. We suggest that these responses are typical of a dynamic photosynthetic apparatus where photosynthetic function is optimized and photooxidative damage is minimized in graduated responses to a combination of nutrients, light quantity and quality.
Figures

References
-
- Bassi R., Soen,S.Y., Frank,G., Zuber,H. and Rochaix,J.D. (1992) Characterization of chlorophyll a/b proteins of photosystem I from Chlamydomonas reinhardtii. J. Biol. Chem., 267, 25714–25721. - PubMed
-
- Bennoun P. and Delepelaire,P. (1982) Isolation of photosynthesis mutants in Chlamydomonas. In Edelman,M., Hallick,R.B. and Chua,N.-H. (eds), Methods in Chloroplast Molecular Biology. Elsevier Biomedical Press, Amsterdam, The Netherlands, pp. 25–38.
-
- Bibby T.S., Nield,J. and Barber,J. (2001) Iron deficiency induces the formation of an antenna ring around trimeric photosystem I in cyanobacteria. Nature, 412, 743–745. - PubMed
-
- Boekema E.J., Hifney,A., Yakushevska,A.E., Piotrowski,M., Keegstra,W., Berry,S., Michel,K.P., Pistorius,E.K. and Kruip,J. (2001) A giant chlorophyll–protein complex induced by iron deficiency in cyanobacteria. Nature, 412, 745–748. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
