

Differential acetylation of Tat coordinates its interaction with the co-activators cyclin T1 and PCAF
- PMID: 12486002
- PMCID: PMC139090
- DOI: 10.1093/emboj/cdf669
Differential acetylation of Tat coordinates its interaction with the co-activators cyclin T1 and PCAF
Abstract
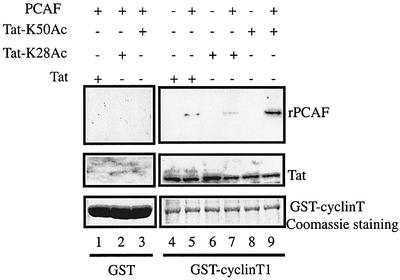
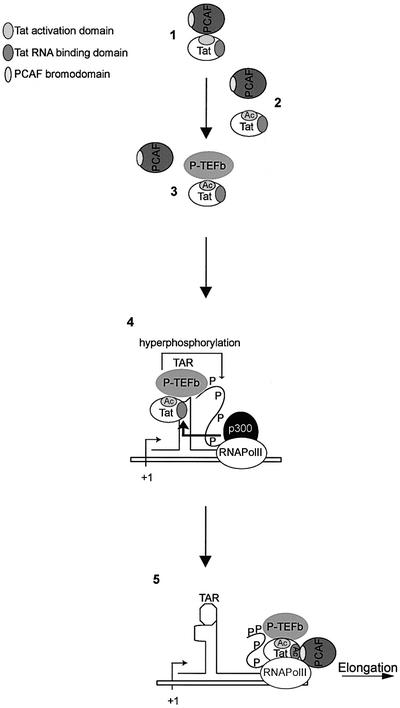
The HIV-1 transactivator protein, Tat, is an atypical transcriptional activator that functions through binding, not to DNA, but to a short leader RNA, TAR. Although details of its functional mechanism are still unknown, emerging findings suggest that Tat serves primarily to adapt co-activator complexes such as p300, PCAF and P-TEFb to the HIV-1 long terminal repeat. Hence, an understanding of how Tat interacts with these cofactors is crucial. It has recently been shown that acetylation at a single lysine, residue 50, regulated the association of Tat with PCAF. Here, we report that in the absence of Tat acetylation, PCAF binds to amino acids 20-40 within Tat. Interestingly, acetylation of Tat at Lys28 abrogates Tat-PCAF interaction. Acetylation at Lys50 creates a new site for binding to PCAF and dictates the formation of a ternary complex of Tat-PCAF-P-TEFb. Thus, differential lysine acetylation of Tat coordinates the interactions with its co-activators, cyclin T1 and PCAF. Our results may help in understanding the ordered recruitment of Tat co-activators to the HIV-1 promoter.
Figures
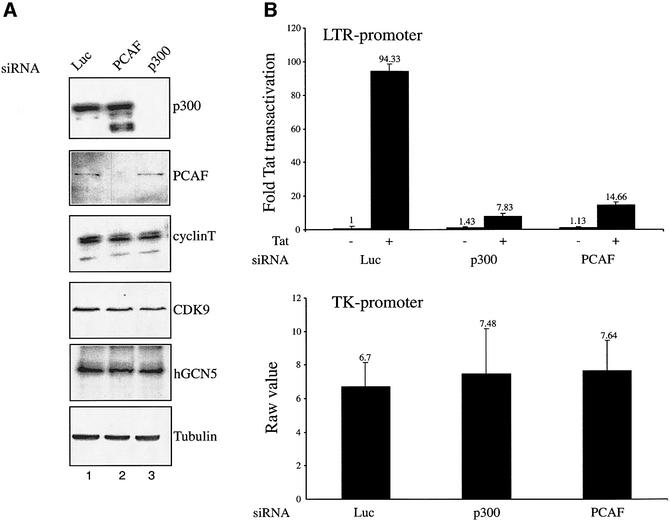
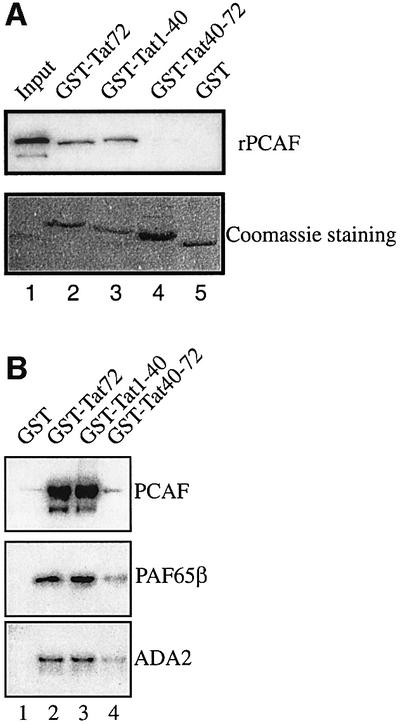
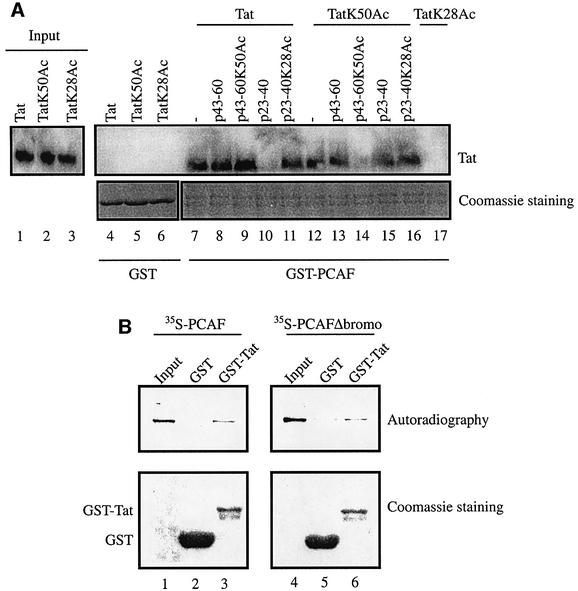
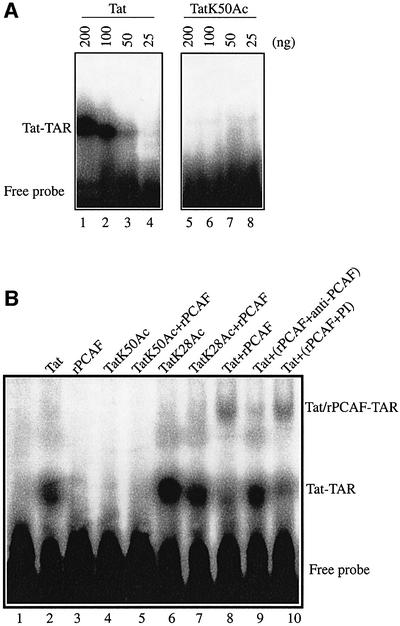
References
-
- Ait-Si-Ali S. et al. (1998) Histone acetyltransferase activity of CBP is controlled by cycle-dependent kinases and oncoprotein E1A. Nature, 396, 184–186. - PubMed
-
- Barany G. and Merrifield,R.B. (1979) The Peptide. In Gross,E. and Meinhofer,J. (eds), Solid phase peptide synthesis, Vol. 2. Academic Press, New York, NY, pp. 1–284.
-
- Beckett D. (2001) Regulated assembly of transcription factors and control of transcription initiation. J. Mol. Biol., 314, 335–352. - PubMed
-
- Benkirane M., Chun,R.F., Xiao,H., Ogryzko,V.V., Howard,B.H., Nakatani,Y. and Jeang,K.T. (1998) Activation of integrated provirus requires histone acetyltransferase. p300 and P/CAF are coactivators for HIV-1 Tat. J. Biol. Chem., 273, 24898–24905. - PubMed
-
- Berkhout B., Silverman,R.H. and Jeang,K.T. (1989) Tat trans-activates the human immunodeficiency virus through a nascent RNA target. Cell, 59, 273–282. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
