

The DNA chaperone HMGB1 facilitates ACF/CHRAC-dependent nucleosome sliding
- PMID: 12486007
- PMCID: PMC139112
- DOI: 10.1093/emboj/cdf692
The DNA chaperone HMGB1 facilitates ACF/CHRAC-dependent nucleosome sliding
Abstract
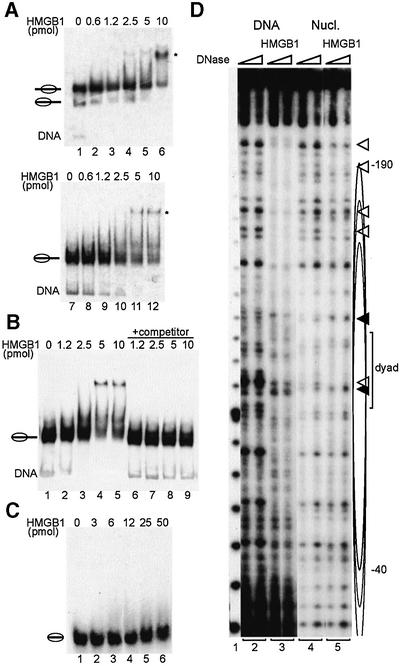
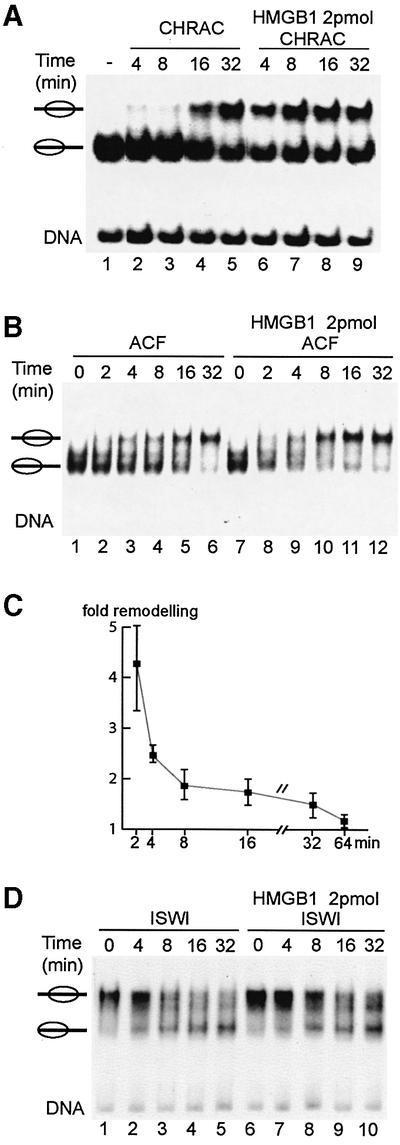
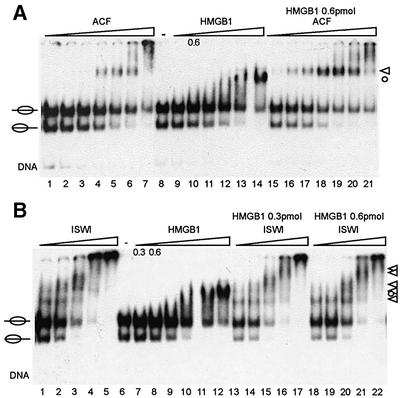
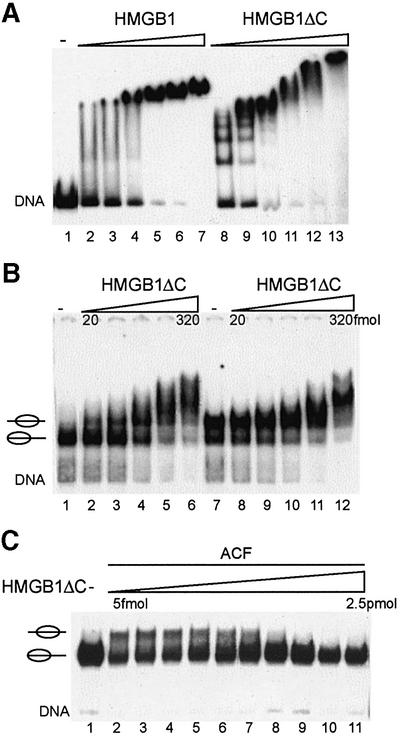
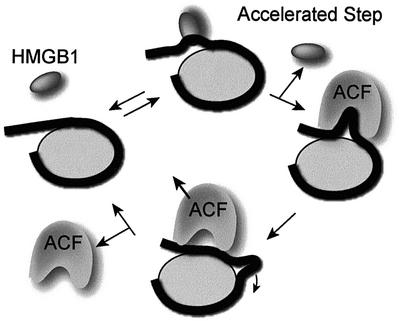
Nucleosome remodelling complexes CHRAC and ACF contribute to chromatin dynamics by converting chemical energy into sliding of histone octamers on DNA. Their shared ATPase subunit ISWI binds DNA at the sites of entry into the nucleosome. A prevalent model assumes that DNA distortions catalysed by ISWI are converted into relocation of DNA relative to a histone octamer. HMGB1, one of the most abundant nuclear non-histone proteins, binds with preference to distorted DNA. We have now found that transient interaction of HMGB1 with nucleosomal linker DNA overlapping ISWI-binding sites enhances the ability of ACF to bind nucleosomal DNA and accelerates the sliding activity of limiting concentrations of remodelling factor. By contrast, an HMGB1 mutant with increased binding affinity was inhibitory. These observations are consistent with a role for HMGB1 as a DNA chaperone facilitating the rate-limiting DNA distortion during nucleosome remodelling.
Figures
References
-
- Aalfs J.D. and Kingston,R.E. (2000) What does ‘chromatin remodeling’ mean? Trends Biochem. Sci., 25, 548–555. - PubMed
-
- An W., van Holde,K. and Zlatanova,J. (1998) The non-histone chromatin protein HMG1 protects linker DNA on the side opposite to that protected by linker histones. J. Biol. Chem., 273, 26289–26291. - PubMed
-
- Beard P. (1978) Mobility of histones on the chromosome of simian virus 40. Cell, 15, 955–967. - PubMed
-
- Becker P.B. and Hörz,W. (2002) ATP-dependent nucleosome remodelling. Annu. Rev. Biochem., 71, 247–273. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
