

Subunit topology of two 20S proteasomes from Haloferax volcanii
- PMID: 12486053
- PMCID: PMC141826
- DOI: 10.1128/JB.185.1.165-174.2003
Subunit topology of two 20S proteasomes from Haloferax volcanii
Abstract
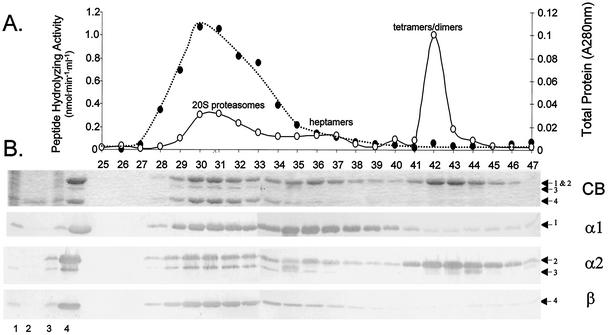
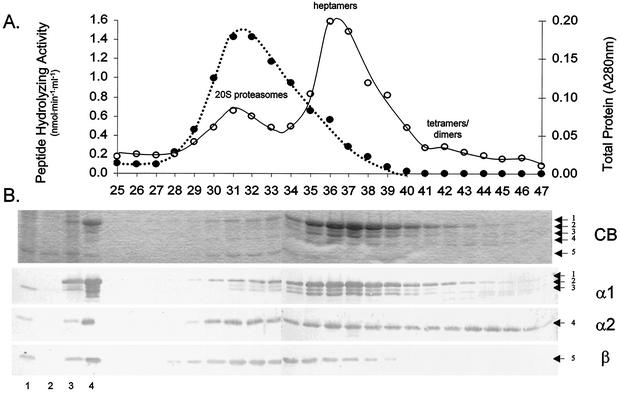
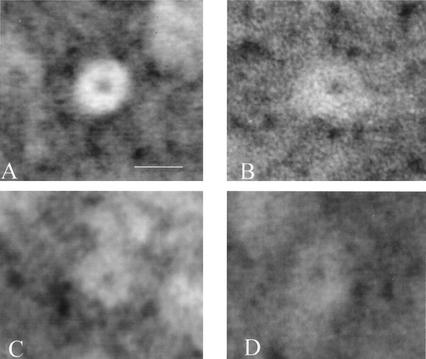
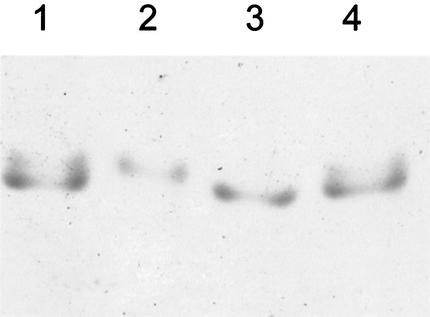
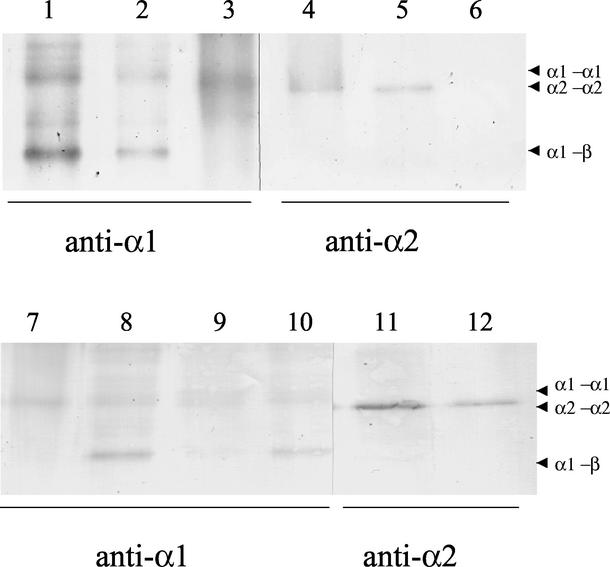
Haloferax volcanii, a halophilic archaeon, synthesizes three different proteins (alpha1, alpha2, and beta) which are classified in the 20S proteasome superfamily. The alpha1 and beta proteins alone form active 20S proteasomes; the role of alpha2, however, is not clear. To address this, alpha2 was synthesized with an epitope tag and purified by affinity chromatography from recombinant H. volcanii. The alpha2 protein copurified with alpha1 and beta in a complex with an overall structure and peptide-hydrolyzing activity comparable to those of the previously described alpha1-beta proteasome. Supplementing buffers with 10 mM CaCl(2) stabilized the halophilic proteasomes in the absence of salt and enabled them to be separated by native gel electrophoresis. This facilitated the discovery that wild-type H. volcanii synthesizes more than one type of 20S proteasome. Two 20S proteasomes, the alpha1-beta and alpha1-alpha2-beta proteasomes, were identified during stationary phase. Cross-linking of these enzymes, coupled with available structural information, suggested that the alpha1-beta proteasome was a symmetrical cylinder with alpha1 rings on each end. In contrast, the alpha1-alpha2-beta proteasome appeared to be asymmetrical with homo-oligomeric alpha1 and alpha2 rings positioned on separate ends. Inter-alpha-subunit contacts were only detected when the ratio of alpha1 to alpha2 was perturbed in the cell using recombinant technology. These results support a model that the ratio of alpha proteins may modulate the composition and subunit topology of 20S proteasomes in the cell.
Figures
References
-
- Akopian, T. N., A. F. Kisselev, and A. L. Goldberg. 1997. Processive degradation of proteins and other catalytic properties of the proteasome from Thermoplasma acidophilum. J. Biol. Chem. 272:1791-1798. - PubMed
-
- Baumeister, W., J. Walz, F. Zühl, and E. Seemüller. 1998. The proteasome: paradigm of a self-compartmentalizing protease. Cell 92:367-380. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
