

Residue R113 is essential for PhoP dimerization and function: a residue buried in the asymmetric PhoP dimer interface determined in the PhoPN three-dimensional crystal structure
- PMID: 12486063
- PMCID: PMC141829
- DOI: 10.1128/JB.185.1.262-273.2003
Residue R113 is essential for PhoP dimerization and function: a residue buried in the asymmetric PhoP dimer interface determined in the PhoPN three-dimensional crystal structure
Abstract
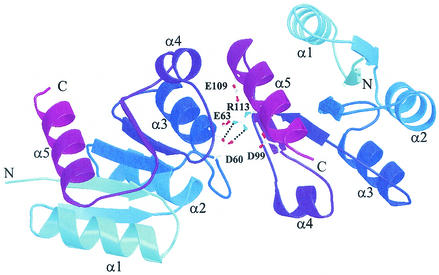
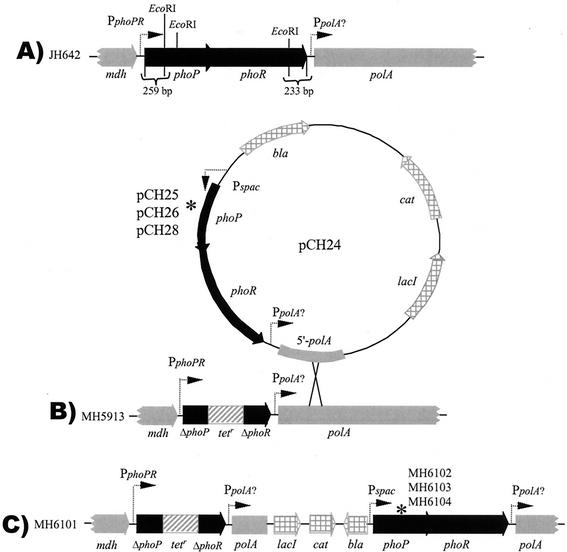
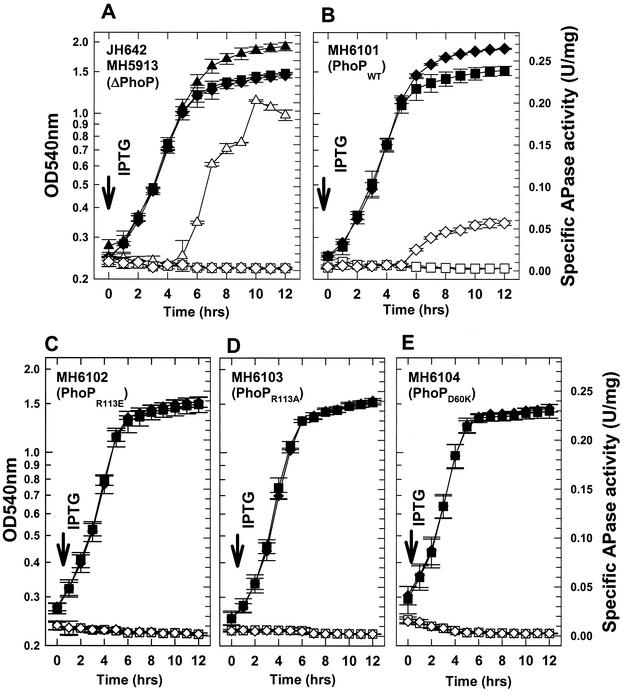
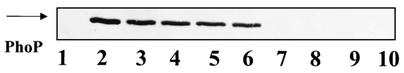
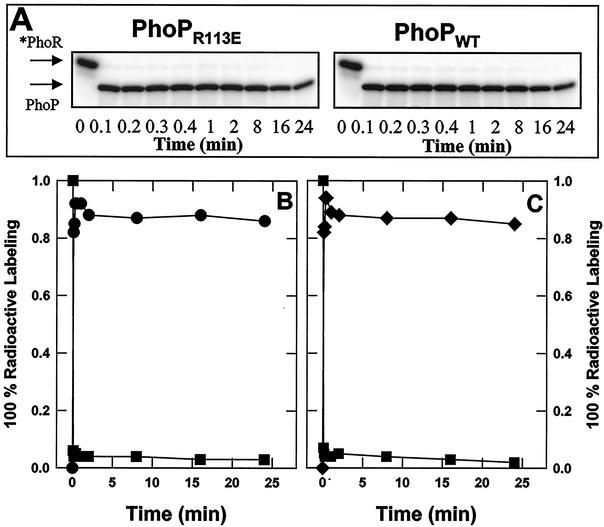
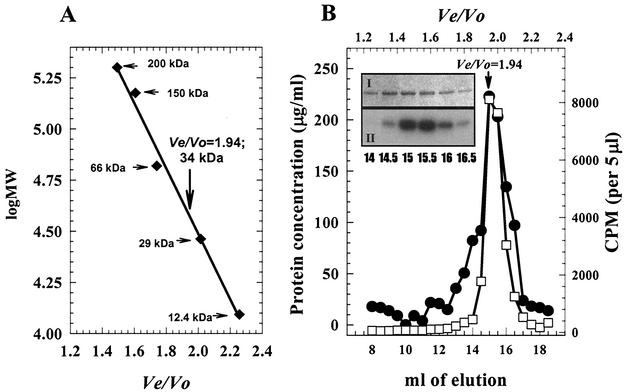
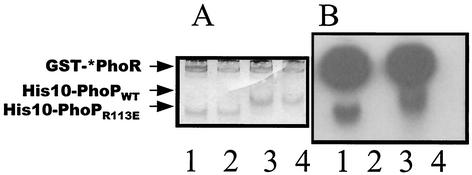
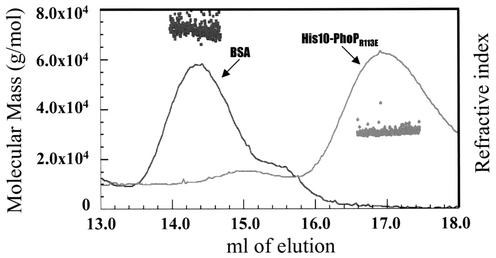
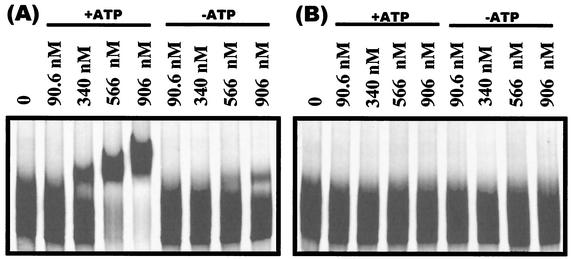
Bacillus subtilis PhoP is a member of the OmpR/PhoB family of response regulators that is directly required for transcriptional activation or repression of Pho regulon genes in conditions under which P(i) is growth limiting. Characterization of the PhoP protein has established that phosphorylation of the protein is not essential for PhoP dimerization or DNA binding but is essential for transcriptional regulation of Pho regulon genes. DNA footprinting studies of PhoP-regulated promoters showed that there was cooperative binding between PhoP dimers at PhoP-activated promoters and/or extensive PhoP oligomerization 3' of PhoP-binding consensus repeats in PhoP-repressed promoters. The crystal structure of PhoPN described in the accompanying paper revealed that the dimer interface between two PhoP monomers involves nonidentical surfaces such that each monomer in a dimer retains a second surface that is available for further oligomerization. A salt bridge between R113 on one monomer and D60 on another monomer was judged to be of major importance in the protein-protein interaction. We describe the consequences of mutation of the PhoP R113 codon to a glutamate or alanine codon and mutation of the PhoP D60 codon to a lysine codon. In vivo expression of either PhoP(R113E), PhoP(R113A), or PhoP(D60K) resulted in a Pho-negative phenotype. In vitro analysis showed that PhoP(R113E) was phosphorylated by PhoR (the cognate histidine kinase) but was unable to dimerize. Monomeric PhoP(R113E) approximately P was deficient in DNA binding, contributing to the PhoP(R113E) in vivo Pho-negative phenotype. While previous studies emphasized that phosphorylation was essential for PhoP function, data reported here indicate that phosphorylation is not sufficient as PhoP dimerization or oligomerization is also essential. Our data support the physiological relevance of the residues of the asymmetric dimer interface in PhoP dimerization and function.
Figures

References
-
- Arantes, O., and D. Lereclus. 1991. Construction of cloning vectors for Bacillus thuringiensis. Gene 108:115-119. - PubMed
-
- Birck, C., L. Mourey, P. Gouet, B. Fabry, J. Schumacher, P. Rousseau, D. Kahn, and J. P. Samama. 1999. Conformational changes induced by phosphorylation of the FixJ receiver domain. Structure Fold Des. 7:1505-1515. - PubMed
-
- Birkey, S. M., W. Liu, X. Zhang, M. F. Duggan, and F. M. Hulett. 1998. Pho signal transduction network reveals direct transcriptional regulation of one two-component system by another two-component regulator: Bacillus subtilis PhoP directly regulates production of ResD. Mol. Microbiol. 30:943-953. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
